Agriculture Reference
In-Depth Information
Induced resistance to
herbivores and necrotrophs
Induced resistance
to biotrophs
-
Mutualistic symbioses
-
-
±
Growth
JA-dependent defences
SA-dependent defences
±
Negate growth retardation
caused by induced resistance
Gene expression
Gene expression
±
Concentration dependent?
SA
JA
Decoy
hypothesis
Activator application
Activator application
PGPR
Mycorrhizal fungi
PGPR
Mycorrhizal fungi
Herbivory/wounding
Necrotrophic pathogen
Biotrophic pathogen
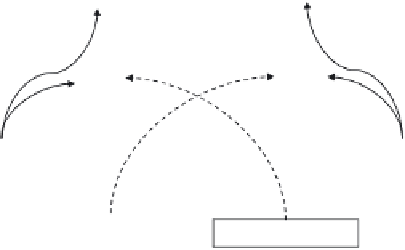
Figure 4.2
Plant resistance against pathogens and insects is regulated by interconnecting pathways
in which salicylic acid (SA) and jasmonic acid (JA) play key signalling roles. The SA pathway is pri-
marily activated in response to biotrophic pathogens whilst the JA-mediated responses are triggered by
necrotrophic infection and insect attack. However, there are no absolutes and recent evidence indicates
that plant defence is more complex than this simple dichotomy suggests. Cross-talk between the SA
and JA pathways results in the activation of distinct sets of defence genes with corresponding trade-offs
between pathogen and insect resistance. This cross-talk is believed to minimise expression of costly
defences that divert resources from plant vegetative growth. Activation of defences in induced resistance
to pathogens may also lead to trade-offs with mutualistic symbioses, for example nodulation or myc-
orrhizal establishment. Note that the use of exogenous elicitors by-passes recognition, although some
have been reported to increase levels of JA or SA in plants. If exogenous elicitors activate induced resis-
tance to both pathogens and insects, competition for limited resources can lead to phenotypically visible
trade-offs.
to suppress SA-induced responses, although reports of this phenomenon are few (Nike
et al.,
1998; Glazebrook
et al.,
2003; Bostock, 2005).
So far, we have dealt with negative effects of interactions between pathogen and insect
resistance. It is important to note therefore, that some workers could fi nd no effect of
induced resistance to pathogens on resistance to insects (Ajlan
& Potter
,
1992; Inbar
et al.,
1998), while there are even reports of positive effects. For example, Stout
et al.
(1999) found that inoculation of tomato leaves with
Pseudomonas syringae
pv.
tomato
induced resistance against both
P. syringae
pv.
tomato
and the corn earworm
Helicoverpa
zea,
while grazing of
Rumex obtusifolius
by the beetle
Gastrophysa viridula
reduced
infection by a number of fungal pathogens (Hatcher & Paul, 2000).
These few examples of crosstalk between the JA and SA signalling pathways highlight
the complex nature of signalling for disease and pest resistance. An interesting insight
into how plants integrate insect- and pathogen-induced signals into specifi c defence
responses was provided by De Vos
et al.
(2005). Using
Arabidopsis,
they tracked the