Biology Reference
In-Depth Information
a growing list of TEs from a increasingly diverse set of sampled organisms. The
rising abundance of mobile element literature from this wealth of data poses a
formidable challenge when attempting a reasonably comprehensive overview of
progress in the area of TE research. We cannot hope, within the scope of this
review, to address the full diversity of elements and their associated biology, or
even to do adequate justice to describing what is known about the major TE
branches. As our own research largely focuses on human TEs, we will primarily
concentrate on progress in our understanding of the human host-element rela-
tionship (Fig. 6.1). Unlike many other higher eukaryotes, humans are currently
bereft of active Class 2 type TEs, although elements of this variety existed
relatively recently (evolutionarily speaking) within our primate lineage (Pace
and Feschotte, 2007). Despite the human and primate-centric focus of this
review, it is of impossible to avoid noting that critical insights into both the
A
TSD
TSD
ORF1
ORF2
~6 kb
AAAAAA
L1 (LINE-1)
TSD
TSD
pol III
AnTACAn
AAAAAA
~300 bp
AB
Alu (SINE)
TSD
TSD
~2-3 kb
Alu-like
VNTR
SINE-R
SVA
B
5
LT R
gag
pol
env
3
LT R
H
ERV
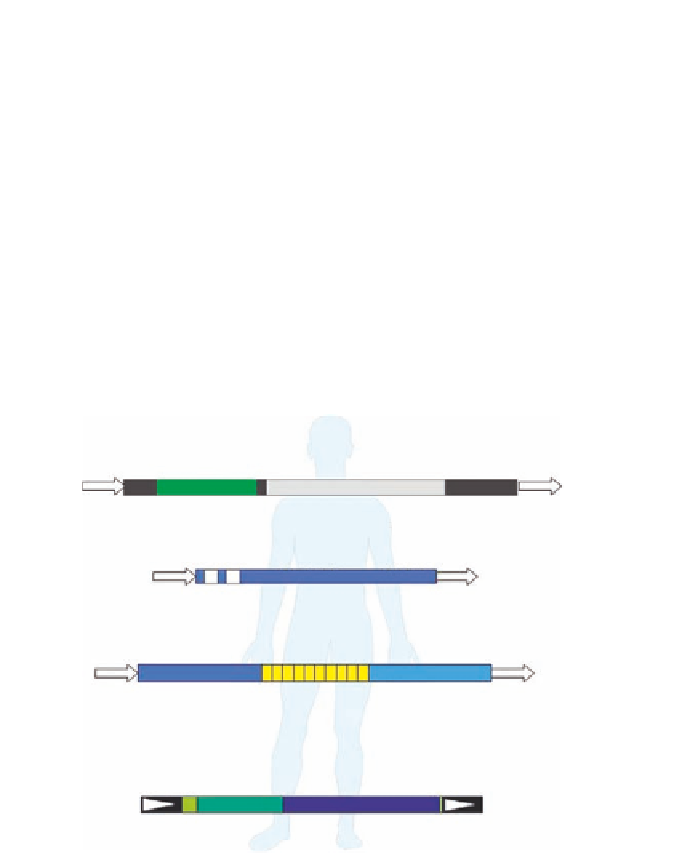
Figure 6.1. The Structure of currently active human TE Lineages. The basic structure of the
currently active human TE lineages is represented. The Pol-II transcribed L1 sequence
contains an internal bidirectional promoter and encodes a bicistronic transcript that
contains two proteins (ORF1 and ORF2). The Alu element consists of two monomeric
units united by an A-rich middle region. The SVA element appears to be a chimeric
construct, derived from components of both SINE and endogenous retrovirus elements.
The structure of HERV elements closely resembles that of the exogenous retroviruses
from which they derived from. They are flanked by two long terminal repeats (LTRs)
and retain protein-coding regions that are recognizably related to viral
gag
,
pol
, and
env
genes.