Agriculture Reference
In-Depth Information
sensor aequorin, which can then sensitively report changes in [Ca
2+
]
cyt
by
giving off light.
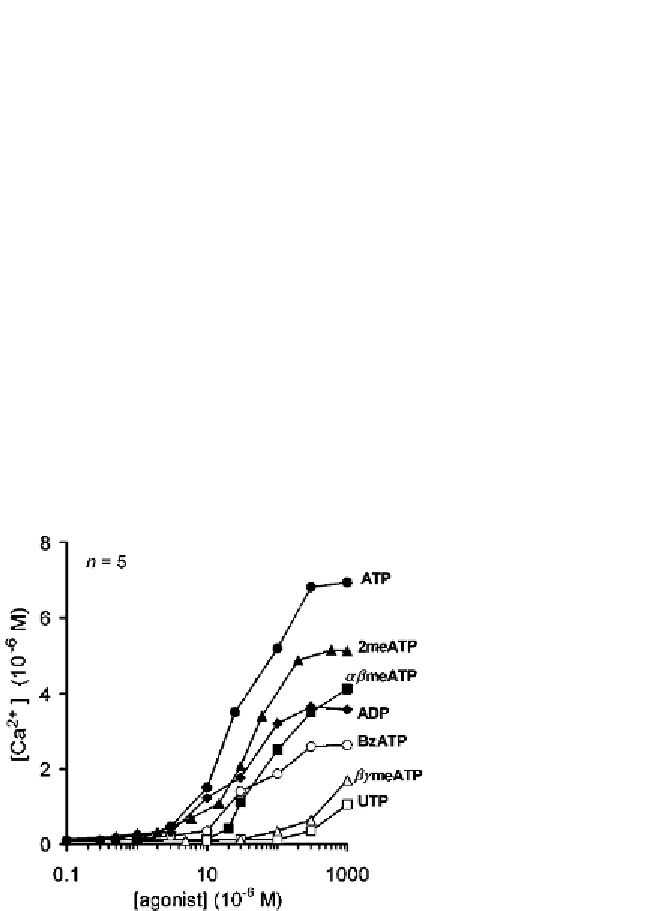
Demidchik et al. (2003) applied nucleotides to excised roots bathed in
a buffered solution containing 10 mM CaCl
2
and found that as little as
300 nM ATP could induce a twofold increase in [Ca
2+
]
cyt
in less than 10 s.
The nonhydrolyzable 2-meATP was almost as effective as ATP, indicating
that agonist hydrolysis was not required for the response (Fig. 15.1). Neither
AMP nor phosphate induced any significant change in [Ca
2+
]
cyt
,andthe
pyrimidine UTP was ineffective below 100 µM, demonstrating the speci-
ficity of the nucleotide action. The P2-receptor antagonist pyridoxalphos-
phate-6-azophenyl-2
,4
-disulfonic acid (PPADS) and the calcium channel
blocker gadolinium, both of which inhibit eATP action at some P2 recep-
tors in animal cells, completely suppressed the response of roots to eATP.
Takentogether,thedoseresponsiveness,substratespecificityandpharma-
cological profile suggested that the response was mediated by a distinct cell
surface receptor, possibly an ion channel. The magnitude of the increase in
[Ca
2+
]
cyt
, from approximately 100 nM to over 400 nM after treatment with
1 µM ATP, was certainly sufficient to turn on calcium-dependent signaling
pathways in plants.
Fig. 15.1.
Externally applied purines and UTP induce an increase in the concentration
of cytoplasmic calcium ions ([Ca
2+
]
cyt
)in
Arabidopsis
roots. Shown are the mean dose-
response curves for effects of different agonists on [Ca
2+
]
cyt
, as estimated by aequorin
luminescence. ATP analogs tested included BzATP, an analog with a modified Rib moiety;
αβ
meATP, which has a methylene insert between the first and second phosphates; and
βγ
meATP, which has a methylene insert between the second and third phosphate. (Taken
from Demidchik et al. 2003, with permission)
Search WWH ::

Custom Search