Biology Reference
In-Depth Information
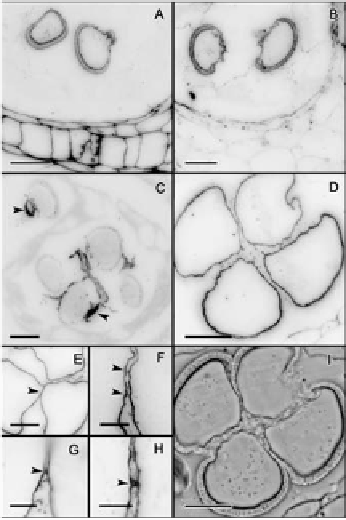
Anti-callose immunofluorescence revealed remnants of callose at the pollen ap-
erture sites where the thick external layer of the microspore wall, the exine, was
extremely thin or absent. These callose remnants were apparent in all microspores
at this stage (Figure 4C). The four microspores showed crosswall cohesion bridges
that stained with antibodies against unesterified and methyl-esterified pectins in
the microspore wall (Figure 4D-H). Following this inter-intine cohesion, addi-
tional deposition of sporopollenin with a joint layering of the four microspores
further strengthened this connection (Figure 4I).
Figure 4.
Establishment of pollen cohesion in Annona cherimola.
(A) Microspore
walls show methyl-esterified pectins, and also (B) unesterified pectins. (C) As callose is digested,
remnants of callose (white arrow) are observed layering the pollen aperture sites. (D-F) Microspores show
crosswall cohesion bridges showing the presence of unesterified pectins. (G-H) Details of crosswall cohesion
bridges, showing the presence of methyl-esterified pectins. (I) Phase contrast of a mature pollen grain showing
internal cohesion and a joint sporopollenin layering. Specific cell components were localized using antibodies
against: methyl-esterified pectin (JIM7) (A, G-H), unesterified pectin (JIM5) (B, D, E, F) Callose (C). A-E, I:
Bar = 10
µ
m. F-H: Bar = 3
µ
m.
Microgametogenesis
As the microspores increased in size, their cytoplasm became vacuolated (Figure
5A) and starch grains were absent (Figure 5B). During this vacuolization, nuclear
migration preceded the first mitosis to form bicellular pollen grains. Following