Environmental Engineering Reference
In-Depth Information
discuss the consequences of inbreeding and genetic
drift for population fi tness and viability and the pros-
pects of 'genetic restoration' (section 7.2.4) in order to
counteract the negative effects of inbreeding and
genetic drift.
tions perform best at their own home site, thus exhibit-
ing local adaptation in the strict sense (Kawecki & Ebert
2004). Also, local adaptation of plants to the biotic
component of their environment is much less evident
than local adaptation to the abiotic environment. In
fact, host-parasite studies, including those of plant-
pathogen and plant-herbivore interactions, generally
show parasite local adaptation (Greischar & Koskella
2007), indicating that host plants suffer more from
their local than from nonlocal biotic threats. This is
especially true for parasites that have higher migration
rates and shorter generation times than their hosts
(Gandon & Michalakis 2002 ). The relative importance
of abiotic and benefi cial or detrimental biotic factors as
determinants of fi tness may thus affect the extent of
local adaptation (Cremieux
et al
. 2008 ).
Nonetheless, local adaptation of populations is, and
should be, an important consideration in the choice of
provenances
used in reinforcement or reintroduc-
tion. For instance, local adaptation may be one of the
main factors explaining why the establishment success
of reintroduced populations of rare or extinct wetland
plant species in Switzerland is best explained by eco-
logical similarity between source and introduction sites
(No ë l
et al
. 2011 ; Figure 7.2 ). Consequently, the choice
of provenances in plant reintroduction programmes is
commonly based on considerations of minimizing geo-
graphical distance to the restoration site or, more
7.2.1
Local adaptation
Local populations have two basic ways in which to
adapt to their environment and changes therein.
Phe-
notypic plasticity
is the ability of an individual to
produce different phenotypes or behaviours from the
same genotype under different environmental condi-
tions, a mechanism that is important for sessile organ-
isms in particular. Although this might suggest that
phenotypic plasticity is a nongenetic mechanism, the
extent to which plants are able to show phenotypic
plasticity in traits ususally has a strong genetic basis
and is, itself, subject to natural selection. The second
mechanism is by
adaptive genetic differentiation
in
response to selection pressures in the local environ-
ment. If the variation in the traits under selection has
a genetic basis, selection may increase the frequency of
alleles in a population that enhance local fi tness. The
latter process will result in
local adaptation
, the
'home advantage' of genotypes in their local environ-
ment, provided that gene fl ow between populations is
restricted, so that the differentiation that evolves is not
continuously swamped by import of alleles that have
experienced different selection pressures (Kawecki &
Ebert 2004). Local adaptation, being an adaptive
response of populations, is of concern in reintroduc-
tion programmes, as translocation implies the risk of
introducing individuals that are maladapted to the
conditions in the restoration area (examples are given
in Chapter 8).
There is ample evidence for local adaptation of plant
populations to their respective environments (Hufford
& Mazer 2003), but it may in fact be less prevalent than
was previously thought. A meta-analysis of plant
transplantation studies (Leimu & Fischer 2008) shows
that in more than 70% of the studied cases local geno-
types outperform foreign genotypes at their site of
origin. However, when these analyses are restricted to
reciprocal transplant studies only, it appears that in
half of the studies in fact one of the two local popula-
tions performed best at both sites (outperforming the
other population both at its local and nonlocal site).
Only in 45% of the cases studied did both local popula-
0.6
0.4
0.2
0.0
-0.2
-0.4
0
0.5
1
1.5
2
2.5
Log (1+Ecological distance)
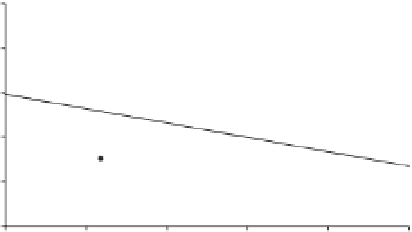
Figure 7.2
The relationship between establishment
success of populations of 14 reintroduced wetland plant
species in Switzerland (log λ, change in log population size
per year) and the ecological distance between source and
introduced site (based on PCA scores capturing differences
in humidity, pH, nutrients, light, temperature and
continentality). Life history traits and propagule
pressure could not predict establishment success.
(From No ë l
et al
. 2011 .)