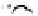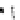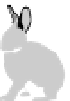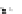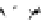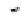Environmental Engineering Reference
In-Depth Information
single herbivore can accelerate species replacement
in favour of stronger competitors. Alternatively, a
herbivore may feed preferentially on stronger com-
petitors, thereby producing a positive indirect effect
on weaker plant species, allowing them to coexist or
even exclude a stronger competitor.
Following Noy-Meir (1975), a number of studies have
stressed the importance of herbivore saturation for the
existence of stable states and threshold effects.
However, multiple stable states can also occur in the
absence of herbivore saturation; that is, at a plant
standing crop where herbivore saturation does not yet
play a role. Van de Koppel
et al.
(1996) have shown
that several herbivore species can co-occur at inter-
mediate standing crop, whereas at high standing
crop plants may become inaccessible for herbivores,
which together results in a hump-shaped pattern
of herbivore density (Fig. 4.5). At low standing crop,
various mechanisms, most notably soil degradation,
may depress plant growth (van de Koppel
et al.
1997)
due to plant-soil feedbacks; a herbivore-induced
decrease in plant standing crop has led to soil degrada-
tion and reduced plant growth. Positive feedback
between reduced plant standing crop and deteriorated
soil conditions has thereby contributed to irreversible
vegetation destruction. It is conceivable that several
mechanisms act simultaneously in many natural
systems. As a result, the growth and consumption
curves intersect twice in a phase plane, even in the
absence of herbivore saturation.
70
(a)
60
50
40
30
20
10
0
120
(b)
100
80
60
40
20
0
70
(c)
60
50
40
30
20
10
0
0
100
200
300
400
Standing crop (g m
-2
)
4.3.2 Plant-parasite interactions
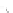
Fig. 4.5
Number (means and individual observation)
of annual droppings of (a) hares, (b) rabbits and
(c) barnacle and brent geese in relation to vegetation
standing crop on the coastal salt marsh of the
Waddensea Island of Schiermonnikoog, The Netherlands.
After van de Koppel
et al.
(1996). Reproduced by
permission of the Ecological Society of America.
Host-parasite interactions can be represented by
resource-consumer models, just as in the case of
predator-prey and herbivore-plant interactions.
A few classic studies, referred to by Zadoks (1987),
showed that pathogenic fungi, selectively parasitizing
plant species in a plant community, may accelerate
vegetation succession. For example, the willow rust
Melampsora bigelowii
killed many seedlings of the
willows
Salix pulchra
and
Salix alexensis
, pioneer
species which formed nearly pure stands on gravel
banks of the River Yukon in Alaska, once the ice had
receded. This might have accelerated succession to birch
and spruce. Another example he referred to is the
massive wane of submarine
Zostera
beds in Dutch
estuaries in the early 1930s, partly due to the pathogen
Labyrinthula macrocystis
. There is increasing interest
in the direct and indirect effects of pathogens and
parasites on the structure of plant communities.
Some recent examples of the roles of pathogens in
determining plant community structure and the con-
sequences of pathogen impact on the functioning