Environmental Engineering Reference
In-Depth Information
The mutual relationship between ecosystem product-
ivity and species richness has been reviewed by
Johnson
et al.
(1996) and Waide
et al.
(1999). In gen-
eral, empirical results have shown a positive, asymp-
totic relationship between ecosystem processes,
measured as productivity and species richness. Only
in a few cases did species richness enhance product-
ivity and stability. These results suggest that once all
functional groups are present, the addition of species
with redundant functions has little effect on ecosys-
tem properties. Reviews of the literature concerning
deserts, boreal forests, tropical forests, lakes and
wetlands lead to the conclusion that extant data are
insufficient to conclusively resolve the relationship
between species richness and primary productivity,
or that patterns are variable with mechanisms equally
varied and complex. For nutrient-related ecosystem
attributes, such as primary productivity, functional
groups will reflect resource-acquisition strategies of
member species. Functional redundancy between spe-
cies is likely to be highest where plasticity or other
mechanisms facilitate coexistence of species that
otherwise compete for consumable resources. However,
careful attention should be paid to Grime's (1998)
observations for plant communities: 'Attribution of
immediate control to dominants does not exclude
subordinates and transients from involvement in the
determination of ecosystem function and sustain-
ability. Both are suspected to play a crucial, if
intermittent, role by influencing the recruitment of
dominants. Some subordinates may act as a filter
influencing regeneration by dominants following
major perturbations.'
Predator-prey interactions are among the most
intensively studied and modelled relationships, prob-
ably because in practice predators and preys are,
in general, easily tangible as individuals, whereas
plants are generally expressed in terms of biomass.
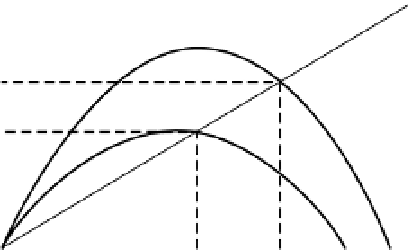
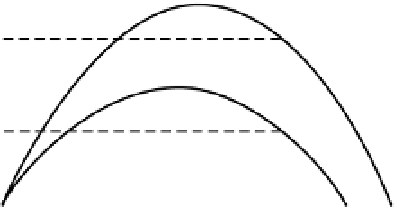
Rosenzweig and McArthur (1963) proposed a hump-
shaped prey isocline in the phase plane. The exact shape
of the prey curve depends on the demographic char-
acteristics of the prey and the carrying capacity of the
environment (Fig. 4.4). Predator numbers should
increase when prey numbers are high, but at high prey
density predators stop increasing because of other
limitations, such as territorial behaviour. Though the
Rosenzweig-McArthur model of predator-prey inter-
actions reveals a wide variety of dynamic behaviours,
Prey zero
isoclines
Predator zero
isocline
(A)
a
b
Prey zero
isoclines
Predator zero
isocline
(B)
a
b
No. of prey (
N
)
Fig. 4.4
Predator-prey isoclines in (A) the classical
Rosenzweig-MacArthur model and (B) the ratio-
dependent model. Two prey isoclines are shown for
more-productive (a) and less-productive (b) habitat.
The equilibrium intersection points are shown by
dotted lines. After Krebs (2001), p. 211. Reproduced
by permission of Pearson Education, Inc.
from stability to strong oscillations, Krebs (2001)
emphasized that all these predator-prey models make
a series of simplifying assumptions about the world,
including a homogeneous world in which there are
no refuges for the prey or different habitats, and the
investigated system is one predator eating one prey.
I mention just two examples showing the complexity
of even a one-prey/one-predator system. Firstly,
Scheiner and Berrigan (1998) measured production and
maintenance costs of plasticity in the freshwater
crustacean
Daphnia pulex
in response to the presence
of chemical signals from a predator, the insect
Chaoborus americanus
. They found scant evidence for
either production or maintenance costs of plasticity,
probably due to a decrease in metabolic rates in the
presence of
Chaoborus
extract, which may have com-
pensated for any cost increases. The second example