Biology Reference
In-Depth Information
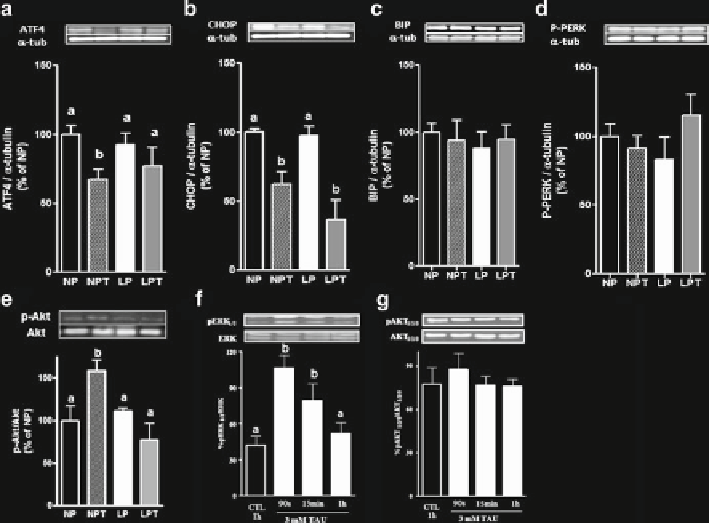
Fig. 14.3
Protein expression of (
a
) CHOP, (
b
) ATF4, (
c
) BIP, (
d
) p-PERK, and (
e
) p-Akt and
a-tubulin (internal control) in islets from NP, NPT, LP, and LPT mice (
n
= 4-7). Groups of fresh
isolated islets from NP mice were incubated with 3 mmol/L TAU for evaluation of (
f
) p-ERK
1/2
/
ERK
1/2
and (
g
) pAkt/Akt ratio. Values are mean ± SEM (
n
= 5); different letters
over bars
indicate
statistical difference;
P
< 0.05 (two-way ANOVA, Newman-Keuls post hoc test)
Ca
2+
handling, since enhanced Ca
2+
uptake and the b2 subunit of the voltage-
sensitive Ca
2+
channel protein expression were observed in islets from TAU-
treated mice (Ribeiro et al.
2009
). Another finding is that TAU supplementation
enhanced intracellular Ca
2+
mobilization using the cholinergic agonist, carbachol
(Ribeiro et al.
2010
), suggesting an increased compartmentalization of the cation
into the ER that may be maintained by SERCA3, since isolated islets from mal-
nourished and control TAU-supplemented rats presented higher expression of
this protein (Batista et al.
2012
) .
Considering these actions of TAU upon intra-reticular Ca
2+
stores and that mater-
nal protein-restriction leads to increased ER stress marker expression in the off-
spring (Sparre et al.
2003
; Vo and Hardy
2012
), we decided to evaluate the expression
of these proteins in pancreatic islets and in the liver from malnourished mice sup-
plemented with TAU. Western blot analysis revealed that malnutrition did not alter
the expression of ER stress markers in pancreatic islets but PERK phosphorylation
and BIP expression were increased in the liver from malnourished mice (Figs.
14.3
and
14.4
). It was reported that young malnourished rats display increased glucose
tolerance and insulin sensitivity (Reis et al.
1997
; da Silva et al.
2012
) , but at 15
Search WWH ::

Custom Search