Biology Reference
In-Depth Information
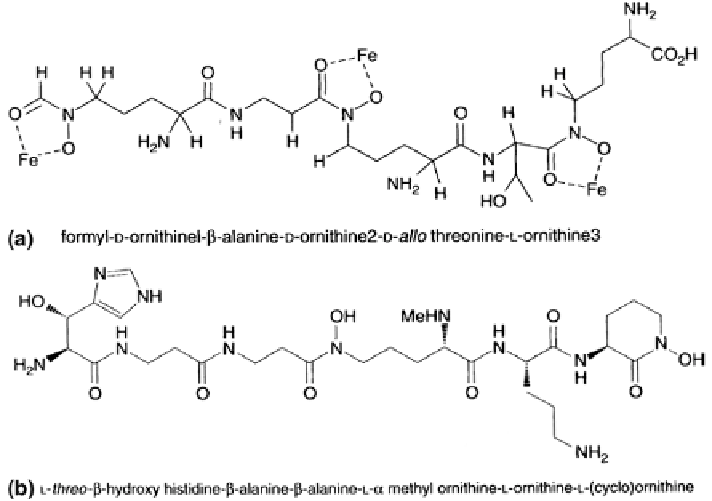
Fig. 2.8
Structures of the exochelins (water-soluble extracellular siderophores) from
a
Mycobacterium smegmatis,
and
b
M. neoaurum
[
94
,
95
]
process and was not inhibited by energy poisons and other agents; it was deduced to
be by a process involving transfer to mycobactin itself [
90
] and could be explained
by the mycobactin preventing a sudden over-load of iron into the cells. Cells that
were iron-deficient had very low contents of porphyrins (Table
2.1
) and thus the key
precursors of haem synthesis were not instantly available to utilize the iron. But even
though iron could not be immediately used and incorporated into cell components,
a mechanism of iron storage was necessary that would then serve as a 'pantry' of
iron. This then began to shape the view that mycobactin was an intracellular store of
iron and that it acquired iron only when there was a sudden availability of it to the
cells (see Fig.
2.11
). Of interest was the finding that an exochelin may be involved
in iron uptake into the leprosy bacillus,
M. leprae
. Somewhat fortuitously, Hall et
al. [
96
] had isolated the exochelin from
M. neoarum
simply because this species
was taxonomically related to
M. vaccae
[
97
] and which, in turn, had been suggested
might be related to
M. leprae
[
98
]. Iron metabolism in
M. neoarum
therefore might
be worth investigating. Richard Hall showed that, by using
55
Fe-labelled exochelin
MN, the iron was taken up by cells of
M. leprae
isolated from armadillo livers but
the process was not one of active transport (as was with its uptake into
M. neoarum
)
and appeared to be by facilitated diffusion [
96
]. The process though was specific in
that there was no transfer of iron when chelated to exochelin MS. However, another
exochelin, this time isolated from an armadillo-derived
Mycobacterium
(ADM)
and which could be grown in the laboratory, also could donate iron to
M. leprae
Search WWH ::

Custom Search