Agriculture Reference
In-Depth Information
300
60
250
50
Aerenchyma
200
40
Wall
(estimated)
150
30
Stele
Total
100
20
50
10
Cortex wall
Cortex
0
0
0
100
200
300
400
500
Distance from apex (mm)
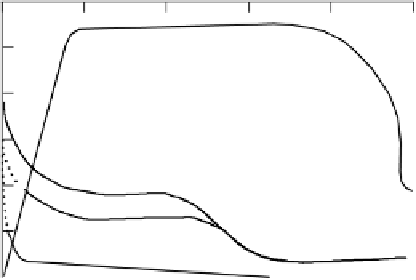
Figure 6.3
Aerenchyma development and changes in respiration rate along the length of
maize roots grown in anoxic media (adapted from Armstrong
et al
., 1991a). Reproduced
by permission of Backhuys publishers
1996). In throughflow systems atmospheric gases are driven or sucked into
the above-ground parts of the plant and then vented from some other point on
the above-ground parts as an O
2
-depleted and CO
2
-enriched exhaust. There are
various possible sources of positive pressure—e.g. humidity-induced diffusion
and thermal transpiration—and of negative pressure—e.g. wind (Venturi forces),
the greater solubility of CO
2
than O
2
(140-fold at 25
◦
C and pH 7), differences in
gas velocities, and thermo-osmosis (references in Jackson and Armstrong, 1999).
Resistance to pressure flow is inversely proportional to the fourth power of the
radius of the conducting vessel, and so large pore-diameters in the diaphragm
partitions of leaf sheath, stem and rhizome are an essential prerequisite for
efficient pressurized flow. A well known example of a pressurized flow system
is the water lily (Dacey, 1980, 1981).
Pressurized flow could in principle occur in a non-throughflow root system,
such as that of rice, driven by dissolution of respiratory CO
2
produced from
gaseous O
2
. However, Beckett
et al
. (1988) have shown that convection by this
means will always be subordinate to diffusion in non-throughflow systems and
will only ever have a minor effect. Hence diffusion is the principle means of
gas transport.
The effectiveness of the internal O
2
transport by diffusion or convection
depends on the physical resistance to movement and on the O
2
demand. The
physical resistance is a function of the cross-sectional area for transport, the
tortuosity of the pore space, and the path length. The O
2
demand is a function
of rates of respiration in root tissues and rates of loss of O
2
to the soil where
it is consumed in chemical and microbial reactions. The O
2
budget of the root
therefore depends on the simultaneous operation of several linked processes and
these have been analysed by mathematical modelling (reviewed by Armstrong