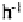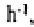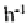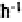Agriculture Reference
In-Depth Information
is known to have existed for approximately 100 years (Watson, 1972) and medium-sized
mounds from a range of species and localities are recorded as having existed for
15-115 years (Grassé, 1984); however, the estimated ages of most mounds are likely
to be towards the lower end of this range.
Bonell
et al.
(1986) measured the rates of erosion of abandoned mounds of the grass
and litter-feeding termite
Amitermes vitiosus
in the semi-arid tropics of northeastern
Australia. Erosion rates were most strongly correlated with high-intensity (>90 mm )
rainfall although lesser intensities also resulted in lower mound erosion rates.
These authors considered it likely that erosion rates would accelerate due to the expo-
sure of internal galleries and the complete breakdown of these mounds through rainfall
was estimated to require
ca.
30 years. A possible 30 years of colony existence may
be added to this to give turnover times of
ca.
60 years for populations of these mounds.
For the soil materials of a population of small mounds (principally soil feeders of
the genus
Cubitermes)
in Zaïre, Aloni and Soyer (1987) estimated a turnover time
of approximately a decade. In addition to erosion by raindrop impact and runoff water,
mounds are also subject to damage by cattle, other large herbivores and by specialised
termitophilous vertebrates, including the birds that create nesting holes in their mounds
(Hindwood, 1959). Over the period of existence of a mound, the total amount of
sediment eroded may substantially exceed the mass of the mature mound, due to the
continuing cycles of erosion and repair of the mound that take place during the period
that the colony occupied it.
The large mounds built by African Macrotermitinae ('high termitaria') exist for
very long periods, possibly many thousands of years. They undergo long periods of
transformation, erosion and increase and develop their own distinctive fauna and flora
(see below), long after the builders of the original mound have died out. Their soils are
often distinctively different from those surrounding them; many have a different texture,
a higher pH and base status (Aloni, 1975; Pullan, 1979) and their organic matter may
differ in its stable C isotopic composition, as shown below.
Galleries and water infiltration
In an arid (350 mm annual rainfall) Australian woodland environment, sub-circular
surface pavements up to 2 m diameter and occupying
ca.
1.6 % of the soil surface area
are the expression of subterranean nests of harvester termites of the genus
Drepanotermes
(Eldridge, 1994). Ponded infiltration rates on the pavement surfaces are very low and
much of the incident water is shed to a surrounding annular area which exhibits
enhanced plant growth with respect to the inter-pavement area more distant from
the mounds. At 113.2 mm ponded infiltration rates in the annular areas are more than
16 times higher than those of the pavement (6.7 mm ) and are slightly higher again
than those of the inter pavement area (103.6 mm ). No differences were apparent in
infiltration under tension and the higher rates of ponded infiltration in the annular zone
were ascribed to the greater densities of termite galleries. In an experimental study
of a desert rangeland ecosystem, Elkins
et al.
(1986) reported higher bulk densities,
lower infiltration rates and higher bedloads in runoff waters from areas in which
subterranean termites had been eliminated by insecticide application.