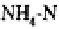Agriculture Reference
In-Depth Information
force in successional processes since they influence the growth and competitive ability
of the plants present at the different seral stages of this process. It has been shown,
for example, that regrowth of
Picea abies
in montane forest of French Alps may be inhibit-
ed by secondary compounds produced by the heath shrub
Vaccinium myrtillum
(André
et al
.,
1987). Allelopathy may also be responsible for the decline of legumes in pastures (Halsall
et
al.,
1995) and for difficulties in the establishment of agroforestry systems that attempt to
combine plantings of trees and herbaceous plants of economic interest (Leigh
et al.,
1995).
Inhibition of nitrification
In chemically-infertile soils, nutrients are mainly conserved within the resistant
soil organic matter fractions and a lack of assimilable C may severely limit microbial
activities (Darici
et al.,
1986). N mineralisation rates may be extremely low in such
diverse ecosystems as natural savannas, artificial pastures or coniferous forests (Scholes
and Sánchez, 1990; Lensi
et al.,
1992; Stienstra
et al.,
1994). Inhibition of nitrification
by water-soluble substances produced by roots has been demonstrated in nutrient-poor
savannas (
e.g.,
Meiklejohn, 1962; Munro, 1966; Lensi
et al
., 1992). Conservation or
the immobilisation of rather than nitrification is an efficient way of conserving N
and preventing C loss from infertile soils (Marrs
et al.,
1991).
3.2.2.3
Inhibition and spread of pathogens by micro-organisms and invertebrates
Soil invertebrates may have a significant role in the suppression of root diseases, through
their direct and indirect effects on the microbial and invertebrate pathogens that affect
roots (Curl, 1988). Dormant fungal propagules freed from fungistasis by root exudates
become able to initiate infection although, when communities of fungal feeding proto-
zoans and nematodes are abundant, these germinands may be consumed before this can
occur (Griffin and Roth, 1979). In soils considered as 'suppressive', infection does not
develop, despite the presence of the root-infecting fungi.
In the case of 'take-all' disease of wheat caused by
Gaeumannomyces graminis tritici,
suppressiveness or reduced severity of disease has been related to the presence of
mycophagous amoebae (Chakraborty
et al
., 1983; Chakraborty and Warcup, 1984); or
lumbricid earthworms (Stephens
et al.,
1994). Other cases of reduced fungal attack have
been related to the activities of mycophagous nematodes, although very high densities
may be necessary to achieve a significant effect (Cayrol
et al
., 1978; Barnes
et al
., 1981).
In certain circumstances, Collembola may also reduce the infection rate of plants
by grazing on pathogenic fungi. Curl
et al.
(1983) showed that the ratio of mycofloral
density in the rhizosphere to non rhizosphere (R/S) diminished from 2.41 to less than
unity (0.41) after the introduction of 500 Collembola into glass tubes
containing 140 g soil and cotton seedlings. Laboratory experiments have demonstrated
that the 'damping-off' disease of cotton seedlings due to fungal infection may be partly
suppressed through the addition of Collembola (Curl, 1988) (Table IV. 12).
Phytoparasitic nematodes may also to be controlled by a number of biological factors
in the rhizosphere. The bacterium
Bacillus penetrans
(Stirling, 1984), the predaceous
fungi (certain zygomycote and other fungi, also protists) including species that trap
nematodes in constricting hyphal loops and against adhesive knobs (Kendrick, 1992)