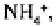Agriculture Reference
In-Depth Information
(v) At a later stage of root growth, the microsite considered occurs in the region of
the root hairs. Since exudation has decreased sharply, bacteria are deprived of nutritional
substrates and either die or return to protected, semi-dormant states. At this stage, the
slowly-increasing populations of micropredators of bacteria are most active. As a result,
part of the nutrients taken up from soil organic matter through priming effects
(
e.g.,
ammonium nitrogen) will be released as metabolites and assimilated by plants.
Micropredation in food chains has been demonstrated to increase nutrient release, and
hence plant production, in microcosm experiments (Ingham, 1981; Anderson
et al.,
1983c;
Setälä, 1990).
3.2.1.2
Roles of mycorrhizae
In nearly all environments, the arbuscular and ectomycorrhizal associations are of most
significance and much of what is discussed below refers to these two types. As indicated
above, dependency on mycorrhizal associations is widespread and the roots of dependent
species often have low surface areas, longer life spans and other properties that differ
from those not so dependent. Mycorrhizal associations generally lead to improved plant
success in both natural and man-influenced environments (Brundrett, 1991; Francis
and Read, 1994) although ineffective and semi-parasitic mycorrhizal associations are
also known.
Perhaps the major effect of the mycorrhizal associations is to increase the volume
of soil explored over that capable of being accessed by the root system alone.
This increase may alter the water relationships of the plant and lead to a greater uptake
of water. The greater soil volume explored also enhances the uptake of such immobile
and slowly diffusing elements as P, K, Cu and Zn. Functional differences occur
between mycorrhizal types and ectomycorrhizal and ericoid mycorrhizal fungi may
access organic forms of N and P that are unavailable to arbuscular mycorrhizal fungi,
and to uncolonised roots (Marschner and Dell, 1994).
The formation of mycorrhizae may also protect the host plant against root diseases
and parasites including bacteria, nematodes and fungi although virus diseases may be
more severe in mycorrhizal than non-mycorrhiza! plants (Perrin, 1990). Protection may
occur in both arbuscular and ectomycorrhizal associations although it is not universal
and may be expected to alter with different combinations of pathogens, mycorrhizal fungi
and hosts. In ectomycorrhizal associations, protection may result through the provision of
a physical barrier between the host plant and the soil environment although it is more
likely that antibiosis and host-plant mediated effects are also involved (Smith and Read,
1997). Interactions between pathogens and potential host plants are strongly influenced
by plant nutrition.
Mycorrhizal associations influence the absorption and uptake of metals that may
occur in potentially toxic concentrations and both increases and decreases in tissue
concentrations have been noted in both arbuscular and ectomycorrhizal associations
(Haselwandter and Bowen, 1996). The fungal associates of ericoid mycorrhizae are
considered to protect their hosts against the potentially toxic concentrations of Al, Zn
and Cu common in their often acid environments (Smith and Read, 1997).
The benefits received by the host plants in mycorrhizal associations are counter-