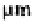Agriculture Reference
In-Depth Information
iii. Less widespread types. Ericoid mycorrhizae are endomycorrhizal associations
formed largely by ascomycote fungi with plants from a number of families within the
Ericales, particularly the Ericaceae and the Epacridaceae and common in heathlands
throughout the world. These fungi form associations with the fine roots of species in the
above families and have significant saprophytic ability consistent with the often infertile
soils within which they and their host plants occur.
Ectendomycorrhizae mainly occur in conifers and have many of the properties of
ectomycorrhizae. They are characterised by the presence of an often-reduced external
fungal sheath and a well-developed Hartig net of internal hyphae. In contrast to the
ectomycorrhizal associations, the internal hyphae of these fungi penetrate the cells of
the host plant. The term ectendomycorrhiza has no functional significance and certain
single fungal strains may form ectencomycorrhizae on one host and ectomycorrhizae
on another (Smith and Read, 1997).
Orchid mycorrhizae. All orchids are achlorophyllous when young and some remain
so throughout life. Orchid seeds are very small - some 50-400 in length (Arditti and
Ghani, 2000) (See also Table I.10) - and the young orchid is completely dependent on
colonisation by a compatible mycorrhizal fungus for development. Adult orchids, both
chlorophyllous and achlorophyllous, usually have mycorrhizal roots or tubers and the
terrestrial forms may be very heavily colonised (Smith and Read, 1997).
While a number of fungi are associated with orchid roots, it is the
Rhizoctonia
species
(Basidiomycota) that are best known. Most fungi are fast-growing saprophytes capable
of using complex polymers as well as more soluble carbohydrates as energy sources.
One species,
Rhizoctonia solani,
is a well-known parasite of commercial crops. The rela-
tionship between the fungus and the orchid may be unstable since in some cases the
fungus may be rejected or become parasitic on its host.
During the heterotrophic stage of orchids that later become chlorophyllous and for
the life of the achlorophyllous species, carbon and other nutrient elements are supplied
by the fungus to the host orchid either from decaying plant materials or through
parasitism in the epiphytic species. It is therefore difficult to understand what benefit
is received by the fungus and these relationships may not be mutualistic.
The geographic distributions of mycorrhizae.
The different types of mycorrhizae are
not distributed at random over the terrestrial surface of the earth. At the biome scale,
different vegetation types are dominated by particular mycorrhizal associations.
Thus, the arctic tundra is dominated by ericoid associations and the northern conifer
forests by ectomycorrhizal associations. Arbuscular associations are particularly
widespread in many tropical rainforests, savannas and other environments although
the tropical areas dominated by the largely ectomycorrhizal Dipterocarpaceae and
Eucalyptus
species (Myrtaceae) are a clear exception to this. Ericoid mycorrhizae
dominate the arctic tundra regions, alpine and other heathlands (Read, 1991).
Symbiotic and free-living N-fixing bacteria
Micro-organisms capable of N fixation are distributed widely between the Eubacteria
and the Archaea and the capacity to fix N has been considered as an ancestral attribute,
although many species have subsequently lost this capacity (Young, 1992).