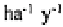Agriculture Reference
In-Depth Information
(Ascomycota) which are able to grow in the absence of a root host, but do not produce
sporocarps (Le Tacon, 1985). Sporocarps are an important dietary item for small
mammals in different parts of the world; the contained spores largely survive passage
through their intestinal tracts and the mammals thereby act as spore vectors for these
fungi (see, for example, Maser
et al.,
1978; Bennett and Baxter, 1989). Annual
production may be substantial (
e.g.,
up to 523 kg in an Australian
Eucalyptus
forest, Johnson, 1994) and ecologically important to wildlife.
Ectomycorrhizae mainly occur in association with trees, largely of the families
Pinaceae, Betulaceae, Fagaceae, Myrtaceae, Dipterocarpaceae and some Leguminosae,
(Newman and Reddell, 1987) together with a number of shrubs and a few herbs. Perhaps
only 3 % of plant species enter into ectomycorrhizal associations but they include
many of the species used in the commercial production of wood (Meyer, 1973) and may
dominate the sites within which they occur (Read, 1991).
ii. Unlike ectomycorrhizae, arbuscular mycorrhizae are internal structures and are
not visible to the naked eye. This form of association is particularly widespread and
involves
ca.
80 % of plant genera, including the agriculturally-important families
Gramineae, Leguminosae, Solanaceae and Compositae. It occurs across a very wide range
of plant taxa including ferns, lycopods, bryophytes as well as the angiosperms and gym-
nosperms. Arbuscular mycorrhizae are particularly common in herbaceous plants but
are also associated with a range of trees and shrubs, including many plant species of eco-
nomic interest, such as
Hevea, Prunus, Citrus, Fraxinus, Acer, Sequoia
and
Podocarpus.
The fungi that form arbuscular mycorrhizal associations are obligate, probably
asexual symbionts with very broad host ranges. The three components of this type of
mycorrhiza are the plant root, the structures internal to the plant root (internal inter- and
intracellular hyphae, arbuscules or organs of interchange and, in some groups, storage
organs, the vesicles) and the network of hyphae (2-27 µ m in diameter, Smith and Read,
1997) that extend beyond the plant root.
The fungal hyphae grow inside the root and penetrate the cell walls (although not
the plasmalemma) to form the microscopic arbuscules which increase the area in contact
with the host. The plant reacts by developing specific fibrillar structures at the point
of contact with the fungus; the cytoplasmic volume is increased, nuclei are enlarged
and starch reserves are reduced (Dexheimer
et al.,
1985). Arbuscules are short-lived
structures and are digested by the host after periods ranging from a few days to a few
weeks. Where vesicles occur, they become filled with lipid droplets during maturation.
Degraded fungal structures appear to be progressively replaced by new ones and the host
may absorb the mineral nutrients released through degradation of the fungal cytoplasm
(Kinden and Brown, 1976).
The fungi associated with these mycorrhizae develop an important network of both
thicker runner hyphae and smaller absorbing hyphae in the soil, although they are less
extensive than those of the ectomycorrhizal association. Estimated field values range
from 0.03 to 13.9 m per gram of soil and up to 46 metres per metre of colonised root,
with higher values obtained from glasshouse studies (Smith and Read, 1997). Such
external hyphae markedly increase the soil volume explored over that which would be
explored by the uncolonised root system (Harley, 1989).