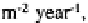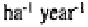Agriculture Reference
In-Depth Information
The microflora is generally divided into bacteria and fungi‚ which perform distinct
functions and often colonise quite separate microsites (see Chapter III.2). Mycorrhizal
fungi have roles that are usually critical to the success of the plants with which they are
associated and are sometimes considered separately. Small populations of predaceous
bacteria and fungi may be found. These are the bacterial predators of other bacteria‚
e.g.‚
of the genus
Bdellovibrio
(Casida‚ 1988) or nematode-trapping fungi (Cooke and
Godfrey‚ 1964; Mitsui‚ 1985; Kendrick‚ 1992). The latter are considered beneficial due
to their potential to control populations of phytoparasitic nematodes. Other fungi of
the genus
Dactyella
have been reported to capture and consume amoebae.
Predatory protists mainly feed on bacteria; cases of mycophagy exist‚ but they are rare
(Petz
et al.‚
1986). They are active consumers: a single heterotrophic nanoflagellate may
ingest three to nine bacteria per hour (Bloem
et al.‚
1989) and Bryant
et al.
(1982)
estimated that 8300 bacteria were consumed for each new amoeba produced in soil
microcosms. Stout and Heal (1967) estimated that protists consume 150-900 g
bacteria equivalent to 15-85 times the standing crop (estimated by the plate
counting technique). Bamforth (1988) distinguished two ecological types of protists:
a group which may serve as food for a large variety of organisms and the others which
are endosymbionts (commensal or parasitic). The latter are regularly found in the gut
of some termites and earthworms (Astomata) and may have endosymbiotic bacteria
within their cytoplasm. Protists of the former group are often considered as transformers
of bacterial protoplasm into higher trophic levels.
Mycophagous amoebae and colpodid ciliates can also ingest fungal spores and may
thereby play a role in the control of populations of phytopathogenic‚ mycorrhizal and
other fungi (Chakraborty‚ 1985; Coûteaux‚ 1985).
Nematodes are another constant component of micro-foodwebs‚ generally one trophic
level above the protists. A few adult nematodes may actually ingest flagellates and some
amoebae (Elliott
et al.‚
1980) although the vast majority are either bacterial-
or fungal-feeders (Chapter III.4.1.2). Nematodes are attracted to their food sources
by a number of stimuli including pheromones‚ and temperature gradients (Freckman‚
1988). Bacterial feeders are active predators that can ingest up to 5000 cells per minute
or 6.5 times their own weight per day when fully active. Overall consumption may be as
high as 800 kg bacteria and the amount of N turned over is in the range of
20-130 kg (Coleman
et al
.‚1984b). This estimate suggests a dramatic role of
microfaunal predation in microbial biomass turnover. Finally‚ mycophagous nematodes
have been reported to suppress plant diseases by ingesting pathogenic fungi (Curl‚ 1988).
The major components of the mesofauna
i.e.‚
Acari‚ Collembola and Enchytraeidae‚
also form part of micro-foodwebs (Moore‚ 1988)‚ although most probably rely more
on mutualistic relationships than predation. Predatory mites can feed on nematodes
(
e.g.‚
Martikainen and Huhta‚ 1990) and have been reported to regulate nematode
density in North American desert ecosystems (Elkins and Whitford‚ 1982). Fungivorous
mites are divided into two groups depending on their growth rates (Moore‚ 1988).
Almost all macrofaunal invertebrates rely on mutualistic relationships with the
microflora for their digestion although some degree of predation exists‚ especially on
protists. For example‚ earthworms can digest ciliates (Piearce and Phillips‚ 1980)
and amoebae (Rouelle‚ 1983). Protists may also participate in the external rumen type of