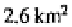Agriculture Reference
In-Depth Information
increase again. The endemic formicine
Formica picea
is found at elevations from 2800
to 4800 m above sea level in the northwest Himalayas (Mani, 1962).
Figure III.57a includes few data from the inter-tropical areas. In a further parallel with
the termites (Section III.4.3.2), substantially higher levels of species richness have been
reported in tropical environments (Hölldobler and Wilson, 1990). At the local level,
these authors cited local richnesses of 172,219 and 272 species from areas of
ca.
in, respectively, Papua New Guinea, Ghana and Peruvian Amazonia. In addition,
Lévieux (1983) reported a total of approximately 230 species from the humid savanna
and gallery forests of Lamto, in the Côte d'Ivoire. Within even smaller areas, a single
tree in Peruvian Amazonia yielded 43 species (Wilson, 1988) while Room (1971) found
128 species in only 250 in a cocoa plantation in Ghana.
Ant species richness is limited by isolation and aridity. A few very isolated areas
(Polynesia east of Tonga, some of the isolated islands of the Atlantic and Indian Oceans)
have no native species of ants (Wilson and Taylor, 1967). Aridity also limits the species
richness of ant communities and dry areas tend to have substantially fewer species
than more mesophytic locations with similar temperature regimes. This was recorded
for African savannas by Lévieux (1983) and for two South American locations by
Kusnezov (1957).
In addition to the species indigenous to particular localities, a number of ant species
have been spread by man to a broad range of countries throughout the world, sometimes
with severe impacts on the local biota. Among these are included the 'tramp' species
which live in close association with man.
4.3.3.6
Spatial distribution of ants
Perhaps the major driving force in ant communities is competition for resources, both
between and within species (Hölldobler and Wilson, 1990). This competition is severe
and unrelenting and results in continuing changes to the composition and community
structure of ant populations and to their spatial dispositions. Competition operates over
a wide spectrum of spatial and temporal scales ranging from the individual colony to
whole faunas and over short and long time scales.
This severe competition has led to niche specialisation and saturation in such species-
rich environments as tropical savannas. It has also led to the horizontal partitioning of the
environment into territories and vertically into a series of partially-overlapping strata.
Territories
The defence of a foraging area containing resources (territoriality) appears to be a general
phenomenon within the ants and may occur between ants of the same or different species
(Hölldobler and Wilson, 1990). It is usually manifested by interference which may
include predation and other competitive interactions leading to the exclusion of other
ants from such resources as food and nesting sites.
In the ants, a territory comprises an area surrounding the nest within which members
of the colony forage for food resources and which is defended against competing
members of the same and other species. Territories may be
absolute
in which case they
are permanently defended against almost all other ant species and very few others