Agriculture Reference
In-Depth Information
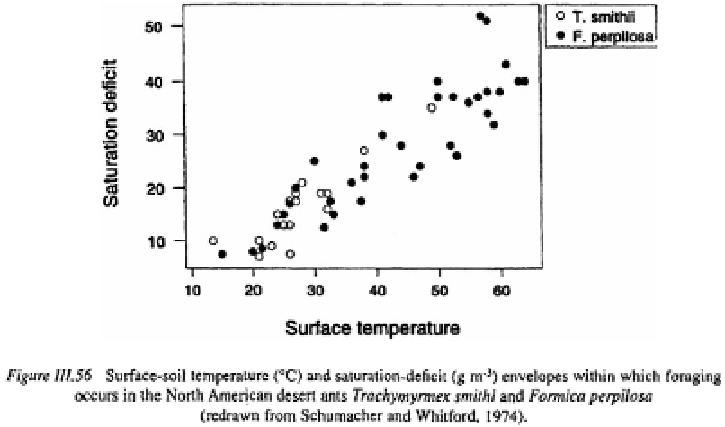
where the rate at which workers returned to a nest of the North American desert fungus
feeding ant,
Trachymyrmex smithi
declined to zero at saturation deficits of greater than
and at soil surface temperatures greater than 50°C. In contrast, workers of
Formica
perpilosa
at the same site continued to forage at temperatures of 55 °C and saturation
deficits of because they were able to replace lost fluid from the honey dew that
they feed on and because of their greater tolerance of water loss (Schumacher and
Whitford, 1974).
Activities are also constrained by strong diurnal rhythms (Hunt, 1974; Roces and
Nunez, 1989) and daily cycles of foraging activity may be modified to avoid predation
and reduce competition (Hölldobler and Wilson, 1990). At a seasonal scale, foraging
may be constrained by environmental conditions. In a Nearctic temperate environment,
the three seed-harvesting species of
Pogonomyrmex
studied by McKay (1981) foraged
most actively during the warmer summer months but were inactive during the winter.
Plant materials
Tobin (1994) argued that while the ants as a group have not specialised in the digestion
of cellulose, their abundance and dominance in many environments is due to their
capacity to exploit plant resources. Many species ingest a wide variety of plant materials,
both vegetative and reproductive, in appreciable and perhaps hitherto unrecognised
quantities (Tobin, 1994). In particular, many ants feed on seeds, seed-associated struc-
tures, plant secretions and other products or little-modified plant materials excreted by
herbivores.
Few ants feed exclusively on plant materials although many use them on a seasonal
or facultative basis. For example, the granivorous ant
Messor pergandei
may consume
leaves, stems and flower parts when seed sources are in short supply (Rissing and
Wheeler, 1976). Vegetative tissues are less commonly eaten although those taken tend