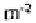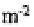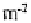Agriculture Reference
In-Depth Information
In most areas, no significant differences in abundance have been observed between
wooded and non-wooded areas. This may be explained by the feeding habits of
Collembola: they do not directly feed on litter, but rather on the soil microflora which is
of comparable abundance in pastures, crops and forest soils. The effects of land manage-
ment are not yet clearly understood. In southern Sweden, Lagerlöf and Andrén (1991)
found densities of 12,430 to 30,410 ind. in four arable crops (fertilised and
unfertilised barley (
Hordeum
sp.), grass and lucerne (
Medicago sativa
) leys) while Persson
and Lohm (1977) recorded much higher values (66,000 ind. in summer, 194,500 in
winter) in a grassland soil. Low soil water potentials may also limit populations.
Biomass generally varies from 80 to 200 mg dry weight (
i.e.,
240-600 mg fresh wt).
The highest values have been recorded in cold temperate ecosystems, notably forests,
where individual mean weights are high. In tropical forests individuals are small and
biomass may be as low as 10-20 mg
4.2.1.2
Community structure
Species structure
The species richness (a diversity) at single sites may be as high as 60 to 80 species in
temperate deciduous forests although large variations occur among environments, with
a clear thermo-latitudinal effect (Petersen and Luxton, 1982). Tundra environments have
low species richnesses with 1-3 species at Signy Island (Antarctica), 7-13 at Devon Island
(Canada), and larger numbers at Barrow (Alaska): 20, Moor House (England): 13-25 and
Hardangervida (Norway): 21-29. In temperate deciduous forests, species richness is in
the range 27-60. In a mixed forest near Paris, 27 species were found over an annual
cycle; nonetheless, only 12 to 19 species were collected at each sampling time (Massoud
et al.,
1984). A maximum of 85 species has been recorded across a range of biotopes
(ß diversity) in mixed forest at Senart in France (Ponge, 1980). In temperate grasslands,
communities may comprise 16-32 species, comparable to the arable land sampled in
Southern Sweden (32 species in cropped plots at Spiboke, Lagerlöf and Andrén, 1991).
No complete list of species seems to be available for tropical environments.
An elevated species richness has also been noted in a number of other soil arthropods
and has been discussed by several authors (see discussion in Section III.4.4.2). High levels
of predation on litter arthropods (up to 70 % mortality in adults, Ernsting and Joose,
1974) and a high mortality due to environmental factors maintain populations at low
densities, with limited, if any, competition (Ponge in Vannier, 1985; Persson, 1980).
Spatial distribution
Collembolan populations are largely confined to the surface holorganic horizons (Petersen
and Luxton, 1982). This illustrates the general function of Collembola as epigeic decom-
posers and, in many sites, 75 to 99 % of the population is concentrated in the upper 2.5 cm.
However, in such ecosystems as the boreal coniferous forest (McLean in Petersen and
Luxton, 1982) or moist African savanna (Athias, 1975), there may also be a significant
proportion of euedaphic populations living below 10 cm depth, down to maximum depths
of 30 or 40 cm. Persson and Lohm (1977) have also observed seasonal variation in
vertical distribution with a marked migration to depth taking place in winter (Figure III.33).