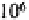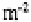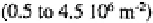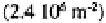Agriculture Reference
In-Depth Information
mull than in moder. Amoebae also have high densities in tundra soils (450-900
individuals at Signy Island (Heal, 1965), and minimal densities in the equatorial dry
forests and savannas of Zaïre and savannas of Ivory Coast
(Coûteaux, 1978). There is some indication that protist abundance and diversity may be
greater in environments with relatively high levels of environmental stress.
Exceptions to this pattern are however known. Persson
et al.
(1980) presented data for
Swedish coniferous forests which clearly suggest that regional and local environmental
variability is at least as important as inter-regional differences in determining the
abundance of protist populations.
4.1.1.2
Community structure
Species richness
Protist communities of temperate and subantarctic communities may comprise
several tens of species in each of the three classes. Many species are cosmopolitan due
to the efficient dissemination of viable cysts and genetic stability (Stout and Heal, 1967).
Consequently, the overall world fauna is unlikely to be greater than a few hundred species.
In New Zealand soils, Stout and Heal (1967) reported species richnesses of 14 to 47
for rhizopods and 21 to 59 for ciliates, with values increasing from grasslands to forests
and subantarctic peats. In England, Stout observed an increase in testacean species from
calcareous mull to acid mor soils (10 to 20) while ciliates exhibited a reverse pattern
(35 to 21). In African savannas, at Lamto (Ivory Coast) 49 species of Testacea have been
found (Coûteaux, 1978) and comparable species richnesses (63) have been recorded in
a variety of forests and savannas in French Guyana (Coûteaux and Chardez, 1981).
Spatial distribution
Because of the nature of their feeding regimes and their water dependence, protists are
particularly concentrated in the upper organic horizons of soils. They are thus
generally more abundant in the litter layers than in the mineral horizons.
Generally, the spatial distribution of protists is similar to that of the micro-organisms
they feed on. This explains why they are such an important component of the rhizosphere;
populations may reach high densities and levels of activity while feeding on the abundant
microbial populations present (see
e.g.,
Clarholm, 1985, Coûteaux
et al
., 1988).
Temporal distribution
In temperate soils, protist populations sometimes show clear seasonal patterns
of variation. Daily counts of populations in English cultivated soils revealed highest
population densities in autumn and lowest in summer (Cutler
et al.,
1922). Similar patterns
have been reported by other authors for temperate soils (Stout and Heal, 1967). Short-term
variation in population numbers is of similar amplitude to that occurring at seasonal scales.
In a pot experiment with a dry arable soil, Clarholm (1981) induced a sudden increase
of bacterial biomass by watering. Populations of bacterial-feeding naked amoebae had
peak biomasses 2 to 3 days after those of the bacteria, and their population density was
increased 30-fold (Figure III.27).