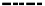Agriculture Reference
In-Depth Information
microsites (under
Cistus
or
Myrtus
shrubs). Nevertheless, the effect of different rhizosphere
microbial populations could not be excluded and the contrasting data of mass loss dynamic
and microbial activity could depend on complex interaction among litter quality, microbial
communities colonizing litters as well as rizhosphere communities.
The decomposition rate of
Fagus sylvatica
litter showed similar results. It was higher in
the beech stand than in a beech-fir stand of Monte Taburno during the first 15 months of
exposition (Figure 7). Its higher decay rate could be related to different microclimatic
condition as well as to interaction among beech, fir and decomposers (Virzo De Santo et al.,
1985). Similar differences were also found by Herlitzius (1983) for beech leaf litter exposed
in beech and
Picea abies
woods.
70
beech-fir
beech
60
50
40
30
20
10
0
0
100
200
300
400
500
600
700
800
900
decomposition time (days)
Figure 7. Accumulated mass loss versus decomposition time of
Fagus sylvatica
litter incubated in a
beech stand and a fir-beech stand on Monte Taburno (Campania Apennines, South Italy). Data from
Virzo De Santo et al., 1985.
M
ICROBIAL
A
CTIVITY
D
URING
D
ECOMPOSITION
Water availability is the main factor affecting microorganism activities in the
Mediterranean area as suggested by the strong correlation between soil and litter water
content and microbial respiration (Papa et al., 2002; Fioretto et al., 1998; 2000; 2001). So,
even if microbial respiration generally increases with temperature, it was at the lowest level in
hot and dry summer and at highest level in spring and autumn for the needle litter of the three
pine types in
P. laricio
wood either as well as for
Q. ilex
litter in the three relative stands and
for
M. communis
and
C.incanus
litters in the low maquis (Fioretto et al., 1998; 2000; 2001 b).
Similar seasonal trend was also evidenced by litter enzyme activities (Figure 8). The
extractable activities of cellulase (EC 3.2.1.4) and xylanase (E.C 3.2.1.8), degrading cellulose
and hemicellulose, respectively, as well as of peroxidase (EC 1.11.1.7), lignolytic enzymes,
showed seasonal variations in
Cistus
,
Myrtus
and
Quercus
litters during the decay process
(Figure 8) (Fioretto et al 2000, 2001 b; Papa, 2000). This pattern was independent from site or
microsite of incubations; therefore, as said before, no significant differences of activity were
found in the
Cistus
litter incubated under
Cistus
and under
Myrtus
shrubs (Fioretto et al.,
2000) as well as among the
Q. ilex
litter incubated in the three woods (Papa, 2000).