Environmental Engineering Reference
In-Depth Information
above). Although these organisms are typically pho-
totrophic, obtaining their nutrients in inorganic form,
other dinoflagellates exhibit some degree of het-
erotrophy. Some are able to ingest food particles by
engulfment of whole cells, others by formation of a
feeding veil (pallium) or use of a feeding tube (phago-
pod). In some cases, these dinoflagellates are pig-
mented and mixotrophic, combining heterotrophic
nutrition with photosynthesis. Other dinoflagellates
such as
Peridiniopsis
are obligate heterotrophs and
colourless. The common freshwater heterotrophic
dinolagellate
Peridiniopsis berolinensis
uses a fine
cytoplasmic filament to make contact with suitable
prey such as insect larvae and then ingests the con-
tents.
Heterotrophic dinoflagellates are particularly
adapted to conditions where photosynthesis is lim-
iting. A number of colourless species, for example,
are found under the ice of frozen lakes during the
winter season, where photosynthesis is severely lim-
ited by low irradiance levels - which are about 1% of
surface insolation.
200
μ
m
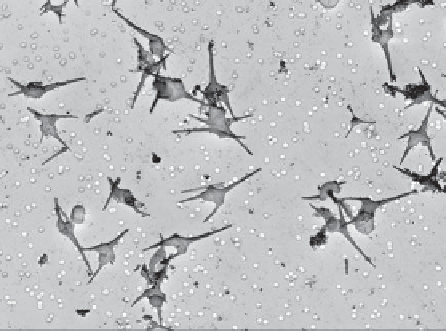
Figure 1.10
Autumn dinoflagellate bloom in a
eutrophic lake. SEM view of epilimnion phytoplank-
ton sample (September), showing almost complete dom-
inance by
Ceratium hirundinella
. See also Figs. 2.8 (sea-
sonal cycle) and 4.56 (live
Ceratium
).
A midsummer to autumn bloom, when phyto-
plankton populations are very high and the surface
watersaredominatedbyeitherdinolagellates(Fig.
1.10) or colonial blue-green algae. At this point in
the seasonal cycle, the surface water concentration
of phosphorus is very low and dinoflagellates such
as
Ceratium
survive by diurnal migration into the
lower part of the lake - where P levels are higher.
Dinoflagellates are adapted to this daily migratory
activity by their strong swimming motion - which
is coupled to an efficient phototactic capacity.
1.8 Cryptomonads
Cryptomonads (Cryptophyta) are a group of rela-
tively inconspicuous algae that are found in both
marine and freshwater environments. They are gen-
erally small- to medium-sized unicells, and in many
standing waters are a relatively minor part of the phy-
toplankton assemblage - both in terms of cell num-
bers and biomass.
An overwintering phase, where dinoflagellates sur-
vive on the sediments as resistant cysts. These non-
flagellate cells (Fig. 2.7) lack an equatorial groove
and initially form in surface waters at the end of
the summer/autumn bloom before sinking to the
bottom of the lake. Germination of cysts occurs
in early summer, either on sediments or recently
mixed waters, giving rise to vegetative cells which
take up newly available phosphorus.
1.8.1 Cytology
The fine structure of cryptomonads is illustrated by
the light microscope images of
Rhodomonas
(Fig.
4.52) and
Cryptomonas
(Fig. 4.53), and by the line
drawing shown in Fig. 1.11. Each cell bears two
unequal flagella, which are about the same length
as the cell and slightly unequal. One of these flag-
ella propels the cell, while the other is stiff and non-
motile. The flagella are inserted near to an anterior
ventral depression, the vestibulum. The cells tend
to be more convex on one side than the other and
Ceratium
and
Peridinium
are geographically
widelydispersed,occurringparticularlyinwatersthat
have high calcium-ion concentrations (hard waters)
and low levels of surface inorganic nutrients (see
Search WWH ::

Custom Search