Biology Reference
In-Depth Information
master
regulator networks share a common network
motif
a fully connected triad, suggesting functional
redundancy across species and cell types. Despite varia-
tions in gene expression in individual cells
[20,41
e
43]
, the
topology and molecular interactions in the core networks
ensure the presence of stable states or attractors encom-
passing the self-renewal state. In this context, attractors are
sets of concentrations of the core transcription factors,
towards which the concentrations evolve over time. In the
case of embryonic stem cells, two or even more attractors
apparently correspond to the pluripotent state
[20,21]
, since
pluripotent cells expressing both high and low concentra-
tions of major pluripotency regulators, such as Nanog, have
been detected
[20,21,44]
. Theoretical studies of network
motifs such as the fully connected triad shown in
Figure 22.1 [45]
, with topology similar to the core pluri-
potency network and few published quantitative models,
largely support the presence of more than one 'self-renewal
attractor' in ESCs
[16,21,22]
.
The described studies allow the narrowing down of the
general question, 'What is self-renewal and pluri-
potency?' to specific questions of the reconstruction of
stem cell regulatory gene networks and identifying and
explaining attractor states for the most critical network
domains.
The core regulatory networks are placed in the context
of much broader arrays of genes, rendering vital stem cell
functions. One such function is susceptibility and response
to external signals.
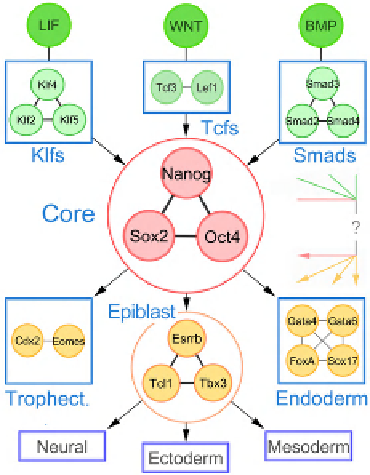
Figure 22.2
summarizes major path-
ways of signaling found in the embryonic pluripotency
gene network. LIF signaling is mediated by members of the
Klf family of transcription factors (Klf4 is essential for
maintenance of induced pluripotent stem cells, iPSC)
[46,47]
. In turn, the expression levels of Klfs are positively
regulated by the core pluripotency factors Nanog, Oct4 and
Sox2
[48]
. The transcriptional repressor Tcf3 represents
another interesting example of interlocking between
extrinsic signaling pathways and the core pluripotency
transcriptional network. A target and negative regulator of
the core pluripotency factors, Tcf3 can also mediate the
Wnt signaling pathway, which has been shown to promote
pluripotency
[49
e
FIGURE 22.2
Signaling pathways in mouse embryonic stem cells.
Links between the major signaling pathways and the core pluripotency
network are shown. Domains with green nodes correspond to 'sensory'
elements, mediating extrinsic signals; domains with yellow nodes are
'executive' elements. Arrows on the right side of the core circuit show
possible feedbacks, bypasses and parallel processing. Direction of links
displays information flow rather than direction of actual interactions
between the genes. Circuits mediating signaling (such as Klfs or Smads,
Gatas) have connectivity features apparently similar to those of the core
pluripotency circuit, which may also serve to achieve functional redun-
dancy at each level of the signaling cascade.
even in the absence of some of the major core transcription
factors, such as Nanog, which highlights the flexibility and
robustness of the pluripotency network
[54]
.
Many network components mediate signaling and
differentiation pathways (
Figure 22.2
) and affect the
dynamic properties of the core network circuit; it is
believed that these inputs, along with the core factors
Nanog, Oct4 and Sox2, contribute to the semi-stable states
described via dynamic attractors (see above). How to
integrate all these inputs, given the high complexity of the
extended pluripotency gene network,
is an important
53]
. In this sense, the position of Tcf3 in
the hierarchy of pluripotency networks resembles that of
Klf4, but it remains to be determined whether Klf4 and
Tcf3 are 'true' integral parts of the (extended) core pluri-
potency circuit. Embryonic stem cells are able to maintain
their pluripotent state even in the absence of external
signals, required for culturing (LIF, BMP) but for a limited
time only
[53]
; external signaling becomes completely
dispensable if cell proliferation suppressors Erk1/2 and Gsk
are inhibited
[22,54]
; in this sense the network is self-
sustainable. Functional redundancy and robustness of the
network are supported by the observation that certain
mouse ESCs can 'adapt' in culture to remain pluripotent
e
emerging problem.
During the early differentiation process the semi-stable
self-renewal states undergo transition and the stem cells
differentiate. One current view is that variation in gene
expression and the consequent existence of attractor states
may be the reason behind this transition. The hypothesis
emphasizes the role of stochastic gene expression in cell fate
determination, so many current models explaining stem cell
behavior include both deterministic and stochastic compo-
nents [18,20
22,42]. Indeed, it has been found that
concentrations of the core factors measured in individual
pluripotent ES cells form statistical distributions featuring
local peaks
e
(bimodality), possibly corresponding to