Biology Reference
In-Depth Information
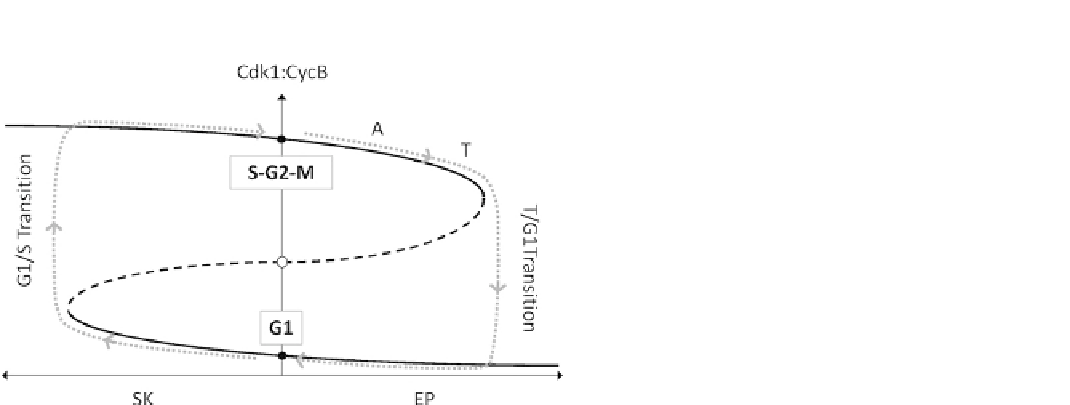
FIGURE 14.6
A dynamic view of cell cycle transitions. The
network in Figure 14.5A creates a bistable switch between a stable
G1 steady state (CycB-kinase activity low) and a stable S-G2-M
steady state (CycB-kinase activity high). In early G1 phase,
SK
z
0 and EP
z
0, and the cell is stuck in the stable G1 state. To
exit G1 phase and begin DNA synthesis, the cell requires a starter
kinase to drive the bistable switch past the saddle-node bifurcation
point and to induce the G1/S transition (gray dotted line). After the
transition is complete, SK activity drops back to zero, but the cell
is now stuck in the stable S-G2-M state. To leave M phase and
return to G1, via anaphase (A) and telophase (T), the cell requires
an exit phosphatase to drive the bistable switch past a different
saddle-node bifurcation point and to induce the T/G1 transition.
After the transition is complete, EP activity drops back to zero, but
the cell is now returned to the stable G1 state.
confirmed experimentally by Uhlmann's group
[50]
; see
Figure 14.7
here.
If we accept this theoretical picture of the G1/S and M/
G1 transitions, then the next logical issues concern regu-
lation of the starter kinase and exit phosphatase.
Start
The G1/S transition in budding yeast is guarded by a check-
point (called 'Start' by yeast physiologists) that controls
production of the starter kinase, Cln2:Cdk1
[51]
. As indicated
in
Figure 14.5
B, Cln2 production is regulated by a transcrip-
tion factor, SBF (a dimer of Swi4 and Swi6), which is kept
inactive by binding to a stoichiometric inhibitor, Whi5
[52,53]
. Phosphorylation of Whi5 and Swi6 leads to activa-
tion of SBF. In
Figure 14.5
Bwe simplify these interactions by
assuming that Whi5 phosphorylation causes dissociation of
the SBF:Whi5 complex. Because Cln2:Cdk1 is one of the
kinases that can phosphorylate Whi5, Cln2 and Whi5 are
involved in a classic double-negative feedback loop that
creates a bistable switch for Cln2-kinase activity. If Cln2:
Cdk1 activity is low, thenWhi5 is unphosphorylated and SBF
is retained in inactive complexes. But if Cln2:Cdk1 activity is
high, thenWhi5 is phosphorylated, SBF is active, and Cln2 is
steadily synthesized.
At the Start transition, this switch is flipped from the
Cln2-low state to the Cln2-high state
[54]
. The Start switch
responds to two crucial physiological signals: cell growth
and mating factor. Newborn daughter cells are too small to
warrant a new round of DNA synthesis
[55]
. They must
grow to a certain critical size before they can pass Start. In
addition, budding yeast cells of mating type
FIGURE 14.7
Reversible exit from mitosis in budding yeast. From
Lopez-Aviles et al.
[50]
; used by permission. In this mutant strain of
budding yeast, the CDC20 gene has been placed under control of
a methionine-repressible promoter (MET-CDC20), and a non-phosphor-
ylable version of Cdh1 protein has been inserted, under control of
a galactose-inducible promoter (GAL-CDH1
CA
; 'CA' for 'constitutively
active'). Finally, a temperature-sensitive allele (cdc16
ts
) of an essential
component of the anaphase promoting complex (APC) replaces the
wildtype gene. This strain is perfectly normal when grown in glucose at
23
o
C (it has Cdc20, endogenous Cdh1, and active Cdc16 proteins). When
grown in glucose
þ
methionine at 23
o
C (time
<
0), these cells arrest in
metaphase, as indicated in the first column of the gel (lots of cyclin B, no
CKI, a small amount of Cdh1
CA
because the GAL promoter is slightly
leaky). Furthermore, the nuclei have a metaphase morphology (micro-
graph at 0 min; red
¼
spindle pole bodies, green
¼
mitotic spindle, blue
¼
DNA). At t
methionine to
induce the synthesis of non-phosphorylable Cdh1 protein, as witnessed by
the third row of the gel. (The fourth row is a loading control.) Because
Cdh1
CA
protein cannot be phosphorylated by the high activity of
CycB:Cdk1 in these cells, the cyclin B subunits are almost completely
degraded by Cdh1
CA
:APC over the course of 50 min (first row of the gel),
and the G1-stabilizing cyclin-dependent kinase inhibitor (CKI) begins to
appear (second row). Furthermore, the nuclei have adopted an interphase
morphology after 50 min of treatment. At t
¼
50 min the cells are trans-
ferred to 37
C to inactivate APC. Despite the fact that the cells appear to
have exited mitosis and returned to G1 phase (low CycB, high CKI), these
cells return to mitosis, as evidenced by the facts that CycB returns, CKI is
degraded, and the nuclei return to metaphase (micrograph at 140 min). If
the treatment is continued for 60 min, and then the cells are transferred to
37
o
C, the cells proceed into G1 phase (not shown; see original paper). This
behavior is clear evidence of a separatrix between two stable steady states:
after 50 min treatment, the cells are still in the domain of attraction of the
stable M-phase steady state, but after 60 min treatment, the cells have
moved into the domain of attraction of the stable G1-phase steady state.
¼
0 the cells are transferred to galactose
þ
a
respond to
pheromone (
factor) by arresting in G1 phase before Start
[51]
. Hence, cell growth promotes the Start transition,
whereas
a
factor inhibits it. Both signals appear to operate
through the activity of Cln3-dependent kinase (and
a second protein, Bck2
[56]
, that is still poorly character-
ized). Cell growth increases the net activity of Cln3:Cdk1
in the G1 nucleus of budding yeast cells, whereas
a
a
factor