Biology Reference
In-Depth Information
Real chemotactic fields usually involve a source that produces chemoattractant continu-
ously (or at least over a time that is long compared to the response time of migrating cells)
and a sink that destroys it. The sink may be localized, sometimes even in the responding cells
because they internalize and destroy the ligand on binding. More commonly, though, it is
distributed, for example as an enzyme activity scattered homogenously throughout the field.
For some unstable molecules, a gradient might be made with no specific enzymatic degrad-
ation. The rate of production (P) and the degradation constant (k
deg
) are important constants
in calculation of steady state gradients in biological situations. Gradients in real tissues are
rarely measured quantitatively, but they are likely to be very complex; in the context of an
embryo, they may not in fact operate in the steady state mode, cells instead responding to
a still-evolving gradient and irrevocable changes being made before steady state is ever
reached. This is especially true when changing embryonic anatomies keep altering the
boundary conditions. It is useful, however, to consider the parameters that shape a gradient
in a very simple model space because the insights from this exercise will be important to the
discussion of gradient sensing later in this chapter.
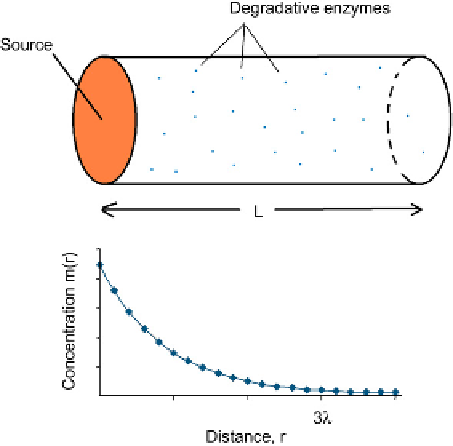
Consider a cylinder of tissue of length L, one end of which is a thin disc of cells that acts as
a uniform source of chemoattractant (
Figure 9.1
). Assume that degradative enzymes are
distributed evenly throughout the tissue and that its sides are impermeable so that overall
diffusion is simply along the cylinder's axis. For such a simple system, the solution to the
diffusion equation when the system has reached a steady state is:
1
p
k
deg
L
l
cosh
fð
L
r
Þ=lg
sinh
L
l
m
ð
r
Þ¼
FIGURE 9.1
Diffusion from a local source in a simple system.