Biology Reference
In-Depth Information
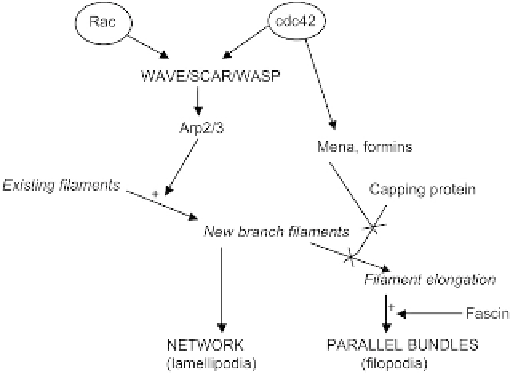
The choice between lamellipodial and filopodial organization therefore depends critically
on the concentration of capping protein. The formation of proper filopodia depends on exten-
sion of filaments taking place at a few discrete sites rather than being spaced out all around
the periphery of the lamellipodium. This may arise if the edge of the cell is inhomogenous,
perhaps having locally high concentrations of molecules such as Ena-VASP and formins, both
of which can bind barbed ends and shield them from being capped while still allowing them
to elongate.
23,24
Such inhomogeneities may be the result of extracellular signals (see Chapters
9 and 11). There may be some degree of self-organization too, though. Consider a cell with
a homogenous distribution of Ena-VASP; once filaments have associated with it and have
started to form
L
-precursors and to extend, they will push the local reserves of Ena-VASP
out of the bulk cytoplasm, producing a temporary local lowering of the concentration of these
molecules in the vicinity of their 'roots' and thus inhibiting the formation of any more filopo-
dia close by. Thus, some degree of spacing may be inherent in the system, even in the absence
of external cues.
CONT
ROL OF FORMATION OF LAMELLIPODIA/FILOP
ODIA
Formation of both lamellipodia and filopodia requires a supply of the basic raw materials,
such as actin, and also requires the activity of branch-initiating complexes such as ARP2/3.
Arp2/3 is activated mainly by proteins of the WASP/WAVE/SCAR family of proteins,
25
and
WAVE/SCAR are in turn activated by proteins such as Abi, Nap1 (
Kette) and Sra1.
26,27
These proteins are induced to interact with WAVE/SCAR by the activated, GTP-bound
form of the small GTPase, Rac. Thus Rac activation acts as a 'master regulator' of lamellipo-
dium construction, and at least some cells fail to migrate in the absence of Rac activity;
28,29
not surprisingly, murine development fails spectacularly if Rac is deleted from the genome.
30
¼
FIGURE 8.7
A simplified overview of the networks that control actin assembly at the leading edge. Activation
of Arp2/3 and formation of new branch filaments is essential to both lamellipodia and filopodia, but modulation of
filament capping tips the balance between these two morphologies.