Biology Reference
In-Depth Information
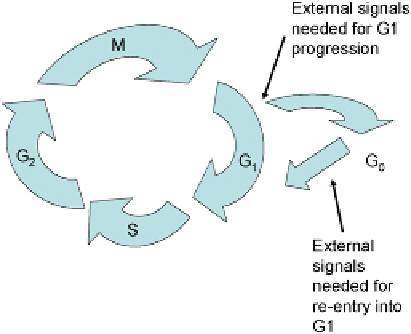
FIGURE 22.5
The basic arrangement of the cell cycle in metazoa. External signals are required at a critical point
of the cell cycle, usually in early G1, in order to ensure cell cycle progression. In their absence, the cell enters the
quiescent state G
0
or dies (Chapter 24). Some cells have their decision point in G
2
instead.
The cell cycles of multicellular animals operate according to broadly similar principles.
The idea of there being many checkpoints is conserved, as is the implementation of them
by cyclins and cyclin-dependent kinases. Indeed, the machinery of specific checkpoints,
such as the metaphase-anaphase transition, is remarkably similar in animals and fission
yeast. What is different, though, is that the cell cycle machinery of animal cells is not self-
sufficient in the way it is in unicells and progress through one critical checkpoint, usually
called the 'restriction point', depends on a combination of signals confirming execution of
the last stage with signals that derive from the outside of the cell.
)
Without both signals
being present, cells enter a quiescent state. For most, the critical checkpoint that depends
on external signals lies in G1 and the quiescent state is called G
o
(
Figure 22.5
). Once cells
have passed the restriction point, they complete their cycle even in the absence of external
signals and only after reaching the decision point again do they enter G
o.
The extracellular
signals that control progress through the restriction point vary between different types of
cells and stages of development and include both positive and negative regulators.
The G1 restriction point of mammalian cells is mediated in part by the actions of an intra-
cellular anti-proliferation protein, Rb. Rb blocks the transcription of genes encoding proteins
that are essential for progression to S phase, such as cyclin A and the enzymes thymidine
kinase and dihydrofolate reductase (both of which are required to make raw ingredients
for DNA synthesis).
19
Rb blocks the transcription of these genes by inhibiting a transcription
factor, E2F, that their transcription requires.
20,21
Rb can itself be inhibited by phosphorylation
by the cyclin-dependent kinases cdk2 and cdk4/6 when they are complexed with cyclin E or
cyclinD1 respectively (
Figure 22.6
). The cdk2/cyclinE and cdk4/cyclinD1complexes are the
indirect target of extracellular signals in a large number of cell types, and a brief illustration
)
Cells in embryos that are still at a stage too early to have shown differentiation into different cell types may
be an exception to this rule.