Biology Reference
In-Depth Information
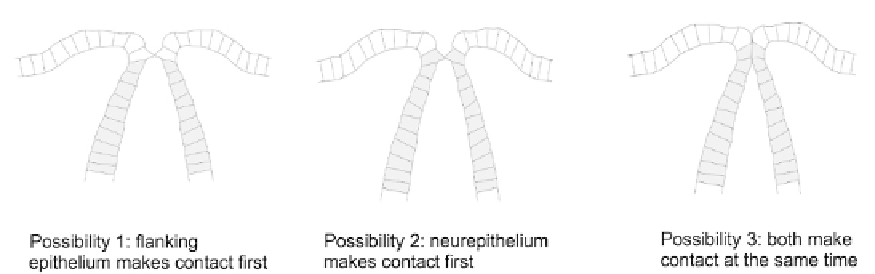
FIGURE 19.10
Different possible orders of contact during neural tube closure.
make contact only later.
26
In each case, cells are already seen to 'reach out' to one another
with fine, highly dynamic projections that are responsible for making the first contact
and inter-digitating to stabilize it.
27
Electron microscopy suggests that full cell-cell junctions
are not made between the projections but the cell adhesion molecule E-cadherin is impor-
tant for reliable closure,
28
suggesting that some kind of adhesive contacts are made. The
apparent absence of clear junctions may be an effect of the general drop in junction produc-
tion that will be described later.
Contact between the converging sides of the neural tube involves signalling as well as
adhesion. In particular, both flanking epithelium and neurepithelium carry Eph-A2 on their
membranes and the neurepithelium also carries Eph-A4. Blocking all Eph-A-type signalling
inhibits fusion
29
without inhibiting any of the events that lead up to it. This strongly suggests
that Ephrin-Eph signalling is required for the sides of the tube to recognize each other's pres-
ence and to rearrange their adhesions.
Rearrangement of adhesions to achieve fusion with new neighbours and schism from old
ones is not at all well understood. It may be very significant, though, that the concentration of
the protein Nf2, which is associated with the assembly of cell-cell junctions near the apical
ends of cells, falls sharply in the crests of the neural folds shortly before fusion.
30
It rises
quickly (perhaps as an indirect result of Eph-A signalling) once cells meet. One possible
reason for this behaviour might be that cells ready to meet one another and to shed old neigh-
bours are kept relatively free of junctions, to be allowed to build them quickly when they have
met their new neighbours. The question of why the new junctions are all with like tissues but
not with unlike tissues, and howwhat must have been apical poles on meeting take on lateral
character, while lateral domains of cells that used to lie on the epithelium-neurepithelium
border take on basal character, remains opaque. There is some information about the
changing expression and location of various molecules associated with apico-basal polarity,
but not enough to make a coherent narrative about cause and effect.
In the first edition of this topic the formation of the penile urethra was given as another
example of fusion following orthogonal invagination, albeit in this case of a progressive
type that took place first at the base of the penis and slowly moved distally. At that time,
this was the consensus understanding of the mechanism and also the consensus explanation
for its very common defect
d
hypospadias (opening of the urethra too proximally). Recent
observation and tracing has, however, greatly changed our understanding of both urethral