Biology Reference
In-Depth Information
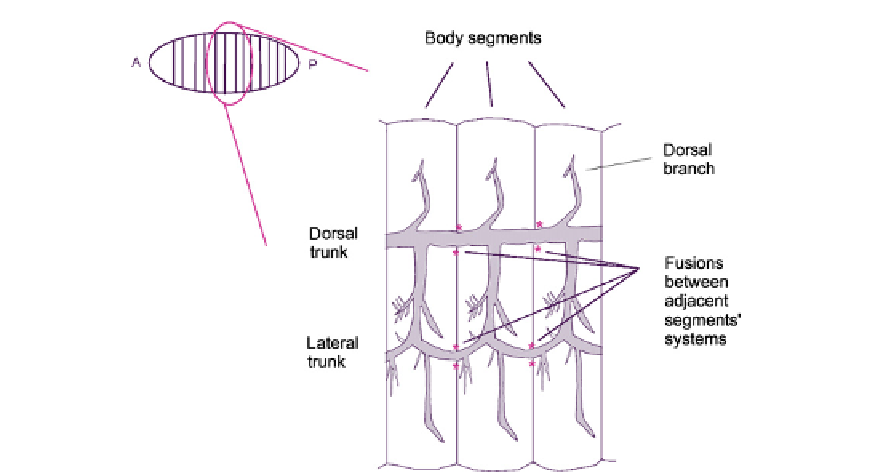
FIGURE 19.3
The connections between tracheal systems of adjacent segments in the embryo of D. melanogaster.
Fusions are flanked by pink asterisks. There is one of these systems on each side of the body.
locations in the inter-segmental walls to form continuous trunk connections leading along the
body (
Figure 19.3
). This probably allows air to circulate, its movement being driven by move-
ments in the body of the insect as it moves.
1,2
Fusion events have been studied in different specific locations within the system and the
results suggest the action of conserved core mechanisms, with some flexibility about the
details. Where branches from the trees of different segments come together to make
the inter-segmental trunks (running horizontally in
Figure 19.3
), the approaching tip cells
migrate on, and are apparently guided by a large bridge cell that makes contact with both
adjacent tracheal systems.
3,4
The first morphological event of tracheal joining seems to be
the production of exploratory filopodia by cells at the tracheal tips. Formation of filopodia
requires the receipt of FGF-type signalling via Branchless and Breathless (Chapter 20) and
depends, as would be expected for filopodia, on activation of the small GTPAse cdc42
(Chapter 8). It also depends on proteins such as Escargot, expressed uniquely in the tip cells.
5
The filopodia project and retract, apparently seeking out a suitable partner
d
a tracheal tip
from an adjacent body segment
d
for fusion.
Once filopodia from approaching tip cells touch, the cell adhesion molecule DE-cadherin
accumulates at the sites of contact (
Figure 19.4
). This accumulation takes about 10 minutes
andmay require newcadherin synthesis, since it is one of the fewaspects of epithelial morpho-
genesis for whichmaternal DE-cadherin RNA is insufficient and zygotically encodedDE-cad-
herin is required.
6
DE-cadherin is normally expressed in the lateral domains of epithelial cells,
mainly near the apex. The filopodia form from the basal sides of the cell, which is the sidemore
typically associated with integrin-mediated cell-matrix adhesion than with cadherin-medi-
ated cell-cell adhesion. The appearance of DE-cadherin in the filopodial contacts is therefore