Biology Reference
In-Depth Information
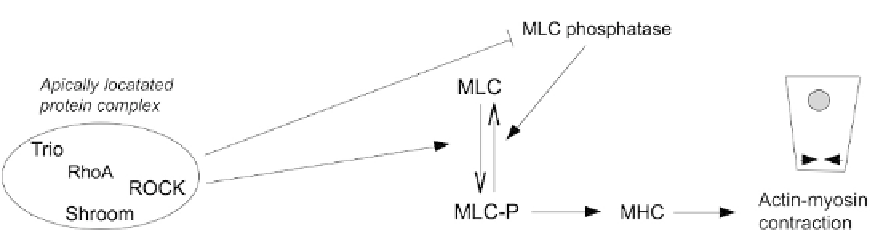
FIGURE 18.4
Control of apical constriction by a complex consisting of apically located RhoA, its activator Trio,
Shroom and ROCK. Activated ROCK phosphorylates, and thereby activates, myosin light chain (MLC) kinase. It
also phosphorylates and inactivates MLC phosphatase, to prevent it from deactivating myosin light chain again.
These combined activates result in the activation of myosin heavy chain and thence apical contraction.
important for constriction. Genetic experiments in invaginating systems in D. melanogaster
also underline the importance of efficient linkage of the actin-myosin system to cell-cell junc-
tions, so that the cytoskeleton has something against which to pull.
6
Cells at many sites of invagination express at least one protein of the Shroom family.
Shroom3 forms a complex with RhoA, ROCK and Trio and, when expressed in MDCK cells,
it can drive apical constriction.
5
At the time of writing, there are conflicting data about
whether Shroom can activate ROCK independently of Rho (Hildebrand
7
suggests it can,
while Plageman et al.
5
state that it cannot). It may be that Rho is needed to target Shroom
apically unless some other system is targeting it to that part of the cell anyway. In the absence
of further information, the consensus control pathway includes a complex with unknown
internal dependencies (
Figure 18.4
). Inhibition of elements of this pathway in real invagi-
nating systems such as the lens pit of the eye (described in more detail below) shows that
invagination fails, or takes place very weakly, in the absence or inactivation of Shroom3,
8
Trio, RhoA, and ROCK.
5,9
Under these circumstances, there is little apical concentration of
activated myosin.
MODELS
FOR AXIAL INVAGINATION (2): MATRIX MEC
HANICS
The sea urchin provides another example of invagination in early embryonic life, in this
case the formation of the archenteron. The epithelium of the vegetal plate flattens and then
invaginates so that a hollow column of epithelium invades the blastocoelic cavity of the
embryo (
Figure 18.5
). The invagination is driven locally and will take place in isolated vegetal
plates, a fact that rules out any mechanisms based on pressure in the blastocoel (in other
words, the archenteron cannot simply be 'sucked in'). It also rules out mechanisms based
on tangential compression in the growing epithelium.
10
There is a local increase in cell divi-
sion in the area around the time of invagination, but treatment of embryos with the DNA
synthesis inhibitor aphidicollin does not prevent invagination from taking place, suggesting
that cell multiplication (Chapter 23) cannot be the primary mechanism.
11
Cells in the vegetal plate show very strong expression of actin/myosin filaments that
run mainly circumferentially under their junctions, and this suggests naturally that the