Biology Reference
In-Depth Information
nature of an epithelium (or an endothelium) has deep implications for morphogenetic mech-
anisms because it prevents cells from undergoing many of the processes typical of mesen-
chymal cells. Instead, the epithelial state allows a different set of morphogenetic processes
that are unavailable to other cell types to occur. To understand how some morphogenetic
mechanisms are restricted while others are allowed, it is first necessary to understand the
structure of a typical epithelium.
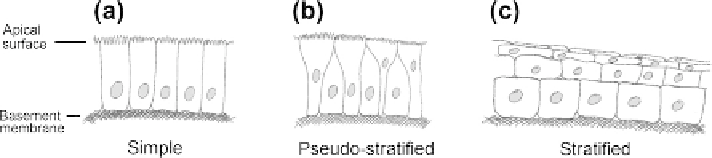
Most epithelia are 'simple' and consist of a single layer of cells attached to a basement
membrane (
Figure 15.2
a). Some, such as skin, are 'stratified' and have several layers that typ-
ically include a basal layer of multiplying cells and overlying layers of differentiating cells
(
Figure 15.2
c). Others are 'pseudo-stratified', with all of their cells making contact with the
basement membrane but arranged so that not all reach the top (
Figure 15.2
b).
Epithelial cells connect to each other and to the underlying basement membrane via
specific types of junctions. In a typical vertebrate epithelium, the junction type found closest
to the apical surface is the tight junction, a complex of proteins that forms an effective seal
between cells so that the fluids bathing the apical domains of cells is kept separate from
that bathing the basolateral domains (
Figure 15.3
). The tight junction also plays a role in pre-
venting the mixing of membrane components across the boundary between the apical and
basal domains.
13
This is important because these domains can possess quite distinct sets of
proteins that arrive by specific targeting of secretory vesicles from the ER/golgi pathway.
Intercellular contacts in the tight junction occur between proteins of the claudin family, which
line up side by side in the membrane to make impermeable belts that surround the cell. Tight
junctions are important for sealing and also originate signals
14
but, in most systems, they do
not seem to play a major role in the mechanics of morphogenesis; they are not, for example,
usually responsible for carrying substantial mechanical forces.
Just below the tight junctions are adherens junctions,
15,16
typically arranged as a 'belt'
running right around the cell. Adherens junctions use cadherinmolecules, usually E-cadherin,
tomediate intercellular adhesion. Their cytoplasmic faces are connected, via adaptor proteins,
to the actin cytoskeleton. This allows tension generated in the cytoskeleton by actin-myosin
contraction to be transmitted between neighbouring cells by means of adherens junctions.
Some of the proteins of the complex on the cytoplasmic face of adherens junctions can trigger
intracellular signalling cascades according to the adhesion state, and perhaps the tension
in, the junction.
17,18
Below the zone of adherens junctions are desmsomes,
19
typically more scattered than are
adherens junctions. Desmosomes also use cadherins, though these are a specialized set (des-
mogleins, desmocollins). Their cytoplasmic faces can also originate signals, and they connect
FIGURE 15.2
The three basic types of epithelium.