Biology Reference
In-Depth Information
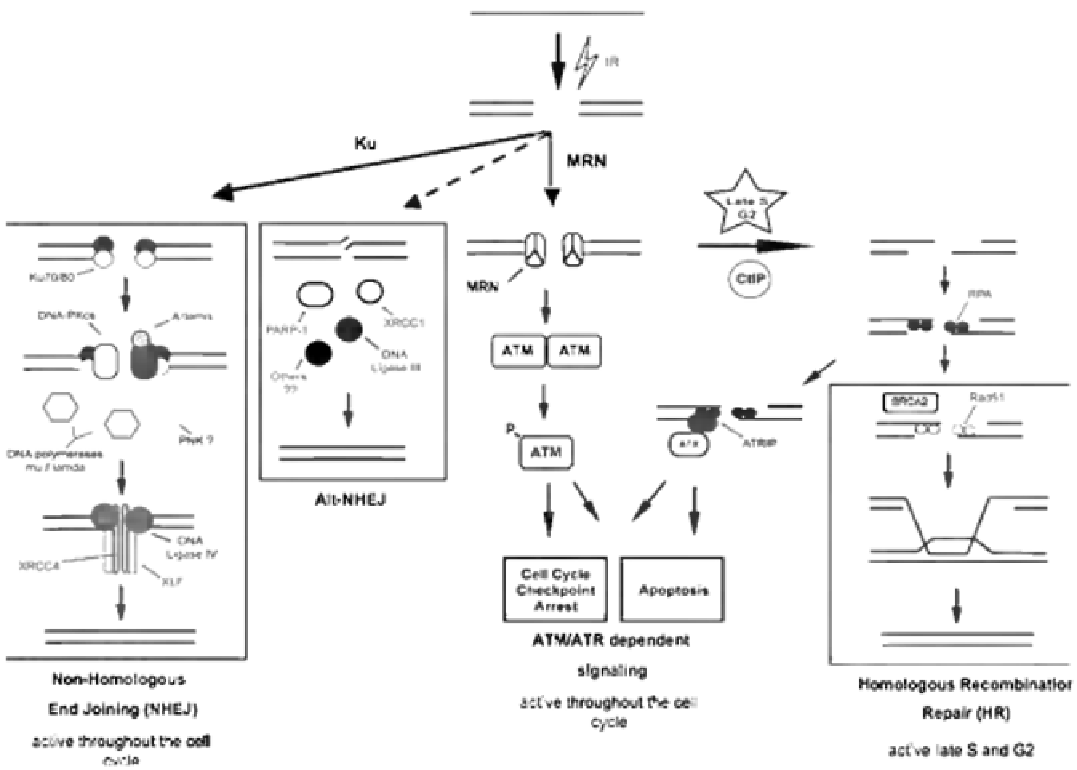
FIGURE 8.5
DNA DSB damage response in mammalian cells. IR induces DSBs, which may be detected by the Ku70/80 heterodimer that
initiates repair by non-homologous end joining (NHEJ) or the Mre11, Rad50, Nbs1 (MRN) complex which directs activation of ATM and
initiation of ATM-dependent signaling pathways (cell cycle independent) as well as activation of DNA resection pathways (in late S and G2)
involving CtIP and other proteins to initiate repair by homologous recombination (HR). The Alt-NHEJ pathway and MRN-dependent activation
of ATR are also shown. How cells choose between these various pathways depends upon cell cycle stage and other, largely unknown factors. See
text for details.
Adapted from Williamson
et al.
, 2009.
290
DNA damage. Central to this DNA damage response is
the ataxia telangiectasia mutated (ATM) protein kinase
(see
Figure 8.5
). Like DNA-PKcs, ATM is a member of
the phosphatidylinositol 3-kinase like (PIKK) family of
serine/threonine protein kinases
78
that phosphorylates
its substrates on serines or threonines that are followed
by glutamines (SQ/TQ motifs).
79
In the absence of
damage, ATM exists in the cell as an inactive dimer.
80
Recruitment of the MRN complex to DSBs triggers the
interaction of ATM with the C-terminus of Nbs1, mono-
merization of dimeric ATM and autophosphorylation on
serine 1981 and other sites, leading to activation of ATM
protein kinase activity.
80,81
In response to DNA damage,
ATM phosphorylates a plethora of downstream targets
including p53, Chk2, H2AX and 53BP1.
82
Indeed, recent
phosphoproteomics studies have revealed that ATM,
and to a lesser extent the other PIKKs ATR (ATM and
Rad3 related) and DNA-PKcs, phosphorylate many
hundreds of proteins in response to DNA damage.
83
e
83
Moreover, there is increasing evidence that ATM and
DNA-PKcs have overlapping and redundant roles, sug-
gesting that their functions are more intimately related
than previously thought.
86
e
88
Non-homologous End Joining (NHEJ)
In the following sections we will describe the mech-
anism of the classical NHEJ pathway, i.e. that involving
Ku, DNA-PKcs, Artemis, XRCC4, DNA ligase IV, and
XRCC4-like factor (XLF, also called Cernunnos), in
the repair of DSBs formed by both exogenous
(primarily IR) and endogenous (primarily V(D)J recom-
bination) sources. Before discussing this pathway in
detail, we will provide a brief historical overview of