Effects of Retroelements on the Host
Copia of Drosophila. As noted, the Ty3/Gypsy lineage is
more closely related to retroviruses than it is to the Copia
In many ways, retroelements are not that different from
lineage (Fig. 6.24). There are also elements in mammals that
extracellular viruses that have learned to infect organisms
resemble LTR retrotransposons, but these may belong to a
and pass from organism to organism. These elements mul-
distinct lineage that arose more recently.
tiply to fill an ecological niche, but are limited to intracellu-
lar spread. However, these retroelements, as noted before,
must be benign in order to avoid being selected against.
Poly(A) Retrotransposons
The ability of retroelements to cause disease appears to
be quite limited, and in many organisms they may cause
Non-LTR-containing retrotransposons, often called
no disease. In at least a few cases, they may even serve a
poly(A)-containing retrotransposons because many have
a poly(A) tract or an A-rich tract at the 3¢ end, are a very
useful function.
The ability of endogenous viruses to cause leukemia in
large family of elements found in virtually all eukaryotes.
AKR mice or mammary tumors in some mice seems to con-
Many of these elements encode RT, and of these many
tradict this observation. However, these diseases are largely
encode Gag-like proteins. The structures of two elements
characteristic of inbred laboratory mice. Wild mice control
from Drosophila melanogaster are illustrated in Fig. 6.25C.
their endogenous viruses much more successfully.
Both encode RT. The I factor element encodes Gag-like
There is no evidence that endogenous viruses ever cause
proteins, whereas R2 encodes proteins that bind nucleic
disease in humans. Humans appear to have no endogenous
acid but are not obviously related to Gag. The elements
proviruses that are ever expressed, although humans do have
that encode RT use it to move by reverse transcription,
defective proviruses that have been present in the germ line
but because they lack LTRs, the mechanism of reverse
for a long time. The absence of endogenous viruses may be
transcription is different.
a function of the long life span of humans. Long-lived ani-
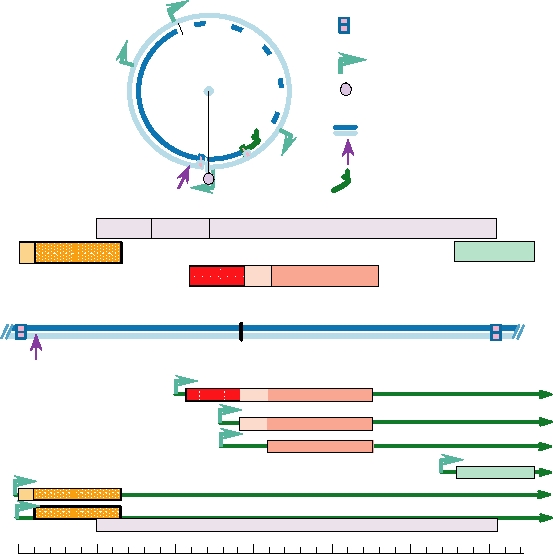
The human genome contains many non-LTR-containing
mals have a longer time span to express endogenous viruses,
retrotransposons. About 20% of the genome, in fact, con-
which would lead to selection against animals that contain
sists of repeated elements called LINEs (long interspersed
such viruses.
nuclear elements, often abbreviated L1). L1 belongs to the
same lineage as the Drosophila elements R2 and I factor
(Fig. 6.24). L1 elements encode Gag-like proteins and RT,
but lack LTRs as well as an envelope gene. It is thought that
FAMILY HEPADNAVIRIDAE
reverse transcription is coincident with integration of L1 into
a new location. In this model, the cell chromosomal DNA is
The hepadnaviruses (hepa from hepatotropic, dna from
nicked by an endonuclease encoded in the retrotransposon,
their DNA genome) share with retroviruses the property
and the nick site is used as a primer for reverse transcription
of encoding RT and replicating via an RNA-to-DNA step.
of the L1 RNA.
They package DNA in the virion, however. The process of
In addition to the L1-like retrotransposons, a simpler
reverse transcription shares features with that described for
class of poly(A) transposons exists that do not encode RT.
the retroviruses but also differs in many important details,
This class includes elements in the human genome called
as described later. Because of the similarities in their mode
SINEs, for short interspersed nuclear elements. SINEs
of replication to that used by retroviruses, the hepadnavi-
include the human ALU sequences present in large numbers
ruses and the plant viruses that replicate via RT are referred
in the genome. It is thought that these elements borrow the
to as pararetroviruses. The hepadnaviruses form a distinct
transcription machinery of the L1-type elements during ret-
taxon in phylogenetic trees, however, and are not particu-
rotransposition.
larly closely related to the plant pararetroviruses or to the
retroviruses (Fig. 6.24). In fact, the plant pararetroviruses,
even though they package DNA in the virion, appear to be
Group II Retrointrons
more retrovirus-like in their replication than are the hepad-
naviruses, consistent with their position in the tree.
Group II introns are self-splicing introns that encode RT.
The hepadnaviruses consist of two genera (Table 6.10).
The structure of such an intron is shown in Fig. 6.25D. These
The genus Orthohepadnavirus contains mammalian viruses
introns are able to move and may have been the source of
and the genus Avihepadnavirus contains viruses of birds.
introns in nuclear genes. They are mostly found in prokary-
The mammalian viruses include hepatitis B virus of humans
otes and in organelles. The RT is translated as a fusion pro-
(HBV), woodchuck hepatitis virus (WHV), ground squirrel
tein from unspliced RNA. Thus, the amount of RT produced,
hepatitis virus (GSHV), and viruses of both Old World and
which determines the ability of the element to move, is regu-
New World primates including woolly monkeys, orangutans,
lated by the efficiency of splicing. The retrointrons group
gorillas, gibbons, and chimpanzees. The primate viruses are
with the retroplasmids and retrons in the RT tree to form a
all closely related and are usually treated as strains of a single
distinct lineage (Fig. 6.24).
TABLE 6.10 Hepadnaviridae a
Virus name
Genus/members
abbreviation
Natural host(s)
Transmission
Disease
Orthohepadnavirus
ACS, hepatitis, cirrhosis, HCCb
Hepatitis B virus
HBV
Humans, chimpanzees,
Horizontal, vertical, IDU,
gibbons,
sexual, blood
Ground squirrel hepatitis B
GSHV
Ground squirrels,
Horizontal, sexual, blood
ACS, hepatitis, HCC
woodchucks, chipmunks
Woodchuck hepatitis B
WHBV
Woodchucks
Horizontal, sexual, blood
ACS, hepatitis, HCC
Woolly monkey hepatitis B
WMHBV
Woolly monkeys
Horizontal, sexual, blood
Hepatitis
Avihepadnavirus
Duck hepatitis B virus
DHBV
Ducks, geese
Predominantly vertical
ACS, hepatitis
Heron hepatitis B virus
HHBV
Herons
Predominantly vertical
a
Hepatitis B virus has a worldwide distribution in humans, as shown in Fig. 6.29.
b
Abbreviations: ACS, asymptomatic carrier state; HCC, hepatocellular carcinoma; IDU, injecting drug users.
Source: Adapted from Fields et al. (1996) Table 1 on p. 2708, with taxonomy according to Fauquet et al. (2005).
virus species with the exception of the woolly monkey virus,
surface antigens or HBsAg. Budding is through internal
which differs by 20% in nucleotide sequence from the other
membranes.
primate viruses and is considered a distinct species. It is rea-
In the description here, the focus is on HBV, but DHBV
sonable to assume that the human virus, which has diverged
has been important for working out the mechanisms of rep-
into a number of strains, arose from one or more of the non-
lication. None of the established cell lines support the com-
human primate viruses. The rodent viruses are distinct from
plete infection cycle of any hepadnavirus, making study of
the primate viruses and diverge by 40% from HBV.
virus replication difficult. Much of what we know comes
The avian viruses include duck hepatitis B virus (DHBV),
from studies of infected liver in experimental animals or
heron hepatitis B virus (HHBV), Ross' Goose hepatitis
studies in explanted primary hepatocytes. DHBV replicates
B virus, snow goose hepatitis B virus, and stork hepatitis
well in primary duck hepatocytes, but the mammalian viruses
B virus. These viruses appear to be widespread and com-
replicate poorly in explanted hepatocytes. Interestingly, a
mon--it is estimated, for example, that up to 50% of free-
number of hepatoma cell lines will support viral replication
living herons in North America may be naturally infected
if transfected with viral DNA. However, attempts to infect
with HHBV. The bird viruses form a distinct lineage. They
them with virus do not result in replication, for reasons that
are closely related to one another but more distantly related
are not clear.
to the mammalian viruses.
As their names imply, all of the known hepadnaviruses
Transcription of the Viral DNA
are hepatotropic, infecting liver cells, and all can cause hep-
atitis in their native host. All have a very narrow host range
A schematic of the life cycle of a hepadnavirus was
that may be determined at least in part by the identity of the
shown in Fig. 1.15. After infection of a cell, the viral nucleo-
receptors used for entry.
capsid is transported to the nucleus. Transport depends on
The hepadnaviral genome is circular and approximately
the phosphorylation of the capsid protein and is mediated
by cellular transport receptors importing α and β. The size
3.2 kb in size, as illustrated in Fig. 6.26A. It consists of
DNA that is mostly, but not completely, double stranded.
of the capsid, about 35 nm, is at the upper limit for transport
One DNA strand, the minus strand, is unit length and has a
through the nuclear pore. The viral genome is uncoated in
protein covalently attached to the 5¢ end, as described later.
the nucleus and converted to a covalently closed, circular,
The other strand, the plus strand, is variable in length, but
dsDNA molecule, called cccDNA. In this process, the pro-
less than unit length, and has an RNA oligonucleotide at its
tein attached to the minus strand is removed, as is the RNA
5¢ end. Thus neither DNA strand is closed and circularity is
oligonucleotide at the 5¢ end of the plus strand, gaps are filled
maintained by cohesive ends.
in, and the ends of the DNA strands are closed. Host repair
Hepadnavirus virions are enveloped and about 42 nm in
enzymes are assumed to carry out this process. The resulting
diameter. The nucleocapsid or core of the virion contains
cccDNA does not integrate into the host genome nor does it
a major core protein called the HBV core antigen, abbre-
replicate as an episome; rather it is maintained as a single
viated HBcAg. The external glycoproteins are called HBV
copy of circular DNA. Note that a primary site of replication
of the virus, and the cells in which most of the studies of
protein may be produced using a non-AUG start codon. As
replication have been conducted, are terminally differenti-
described before, four classes of mRNAs are produced by
ated hepatocytes which divide only rarely and in which there
initiation at the four promoters which form an overlapping
is no ongoing DNA synthesis. Thus, the virus has evolved
set that lead to the production of seven proteins in the mam-
other means for amplification of its genome.
malian viruses and to five or six proteins in the avian viruses
The cccDNA is transcribed by cellular RNA polymer-
(Fig. 6.26B).
ase II to produce several mRNAs (Fig. 6.26B). Only one
Both C and P are translated from the largest (3.5 kb)
strand is transcribed. Four different promoters in the
mRNAs, which are slightly longer than unit length. Two
DNA of the mammalian viruses lead to the production
mRNAs are produced starting at this promoter. One mRNA
of unspliced transcripts of lengths 3.5 kb (i.e., slightly
is slightly longer and is translated to produce the protein
greater than unit length), 2.4, 2.1, and 0.7 kb, all of which
called preC. The shorter form of the 3.5-kb mRNA, which is
terminate at the same poly(A) addition site (purple arrow
also called pgRNA, lacks the AUG used to initiate transla-
in the figure). More than one start point is used in the
tion of preC. A downstream AUG in this mRNA, which is in
case of two of the promoters, and from these two pro-
the same reading frame as preC, is used to initiate translation
moters, RNAs with two different 5¢ ends are transcribed
of the protein called C, which is the major capsid protein
that serve different functions, as illustrated in Fig. 6.26B.
of the virion. PreC has a different fate. It is inserted into
The RNA transcripts are capped and polyadenylated. The
the endoplasmic reticulum during synthesis and transported
polyadenylation signal in the mammalian hepadnavi-
through secretory vesicles, undergoing cleavages to remove
ruses is TATAAA rather than AATAAA, and the use of
an N-terminal signal sequence and some C-terminal resi-
this suboptimal signal appears to require viral sequences
dues. It is secreted from the cell as a 17-kDa protein called
upstream of this site.
HBeAg. HBeAg may be important for the establishment of
Transcription of viral RNA is most efficient in hepato-
a chronic infection in infants (see later). HBcAg, HBsAg,
cytes. At least some of the promoters require transcription
and HBeAg are all used as clinical markers of infection and
factors such as hepatocyte nuclear factor 1, present primarily
virus replication. In general, HBeAg is associated with more
in hepatocytes, for optimal activity. Furthermore, at least
aggressive clinical hepatitis.
two enhancer sequences are known to be present in the DNA
The gene for P is downstream of C (Fig. 6.26B). It is
that function most efficiently in hepatocytes. In addition to
in a different reading frame than C and partially overlaps
the various cellular factors, the X gene product upregulates
C. The mechanism by which translation of P is initiated
transcription of viral DNA.
is not yet resolved. It is translated from the same mRNA
The four classes of mRNAs are exported to the cytoplasm
as is C (i.e., pgRNA), but initiation is internal, using the
for translation and assembly of virions. Export is facilitated
start AUG of P, rather than being produced by some form
by a sequence element of about 500 nucleotides called the
of frameshifting and cleavage. Internal initiation does not
posttranscriptional regulatory element (PRE). This element
appear to use an IRES but appears to be cap-dependent,
is required because the major hepadnaviral RNAs are not
and some form of ribosome scanning has been invoked in
spliced. Thus, PRE is functionally analogous to RRE of HIV
order to position the ribosome at this start site. This process
or CTE of the simple retroviruses, but it is not known if the
is inefficient, and about 200 copies of C are produced for
mechanisms by which these elements effect export are the
each copy of P.
same.
Three forms of S are produced, a long version called L or
preS1, a medium size version called M or preS2, and a short
version S (Fig. 6.26B). These differ only at their N termini
Synthesis of Viral Proteins
and are produced by using different in-frame AUG initiation
Synthesis of the viral proteins is complex. Four genes are
codons. PreS1 is translated from the 2.4-kb mRNA, whereas
usually recognized. The core gene gives rise to two products
preS2 and S are translated from two forms of the 2.1-kb
called precore (preC) and core (also referred to as HBcAg
mRNA in a manner similar to preC and C. As stated, only
or simply C). The polymerase gene gives rise to RT-RNase
the proteins corresponding to L and S are produced by the
H (usually called the polymerase or P). The surface protein
avian viruses.
gene gives rise to three proteins in mammals called preS1,
X is translated from the 0.7-kb mRNA. As described, it
preS2, and S (also called HBsAg), but to two proteins in
is not known if an X protein is produced by the avian hepad-
birds corresponding to preS1 and S. The X gene gives rise
naviruses.
to a protein called X, so called because its function was
The viral genome is very compact (Fig. 6.26). Over half
originally unknown and its complete range of functions is
of it is translated in two reading frames. The P gene requires
still obscure. The X protein has been long considered to be
about three-quarters of the coding capacity of the genome
produced only by the mammalian viruses, but a region cor-
and this gene overlaps each of the other three genes. The S
responding to X is present in the avian viruses and an X
gene is completely contained within the P gene.
S2
A.
Genome organization
Repeated Elements (DR1 and
Eco RI
DR2)
S1
Transcriptional promoters
P Protein on 5 end of
nt 1 nt 3221
minus strand DNA
dsDNA
X
Polyadenylation site
Oligoribonucleotide
B.
C
Open reading frames
Frame 3
TP (spacer)
POLYMERASE
RNAse H
Frame 2
Protein X
Pre
CORE
Frame 1
PreS1
S
PreS2
cccDNA
DR1
Eco RI
DR2
(+) DNA
() DNA
C.
Transcription and translation of m RNAs
S1
An
preS1 RNA
S2
An
preS2 RNA
S2
An
S RNA
X
An
X RNA
C
preC RNA
An
C
C RNA
An
(or pg RNA)
Nucleotides
0
500
1000
1500
2000
2500
3000
FIGURE 6.26 Genome organization of hepatitis B virus. (A) Circular map of the genome showing the locations of the
repeated elements (DR1 and DR2), the four known promoters (blue-green arrows), and the gap of variable length in the plus
strand of the DNA. This map is numbered from the beginning of the pgRNA. Some authors number the nucleotides from the
unique Eco RI site (here at nt 1407). (B) Linearized map of cccDNA showing the open reading frames. Below this are shown
the six mRNAs and the proteins (colored blocks) translated from them. Note that the pC RNA and C RNA (pg RNA) are of
more than genome length, and that all mRNAs end at the polyadenylation site (purple arrow) at nt 121 (but have been shown
extended to the right for clarity). (C) Transcription and translation of mRNAs. All transcripts are made from the (-)DNA (light
blue) as template, in a clockwise direction on the circular map, or left to right in the linearized map. Data for constructing this
diagram come from Yen (1998), Hu and Seeger (1997), and Fields et al. (1996) p. 2706, and details provided by Dr. James Ou.
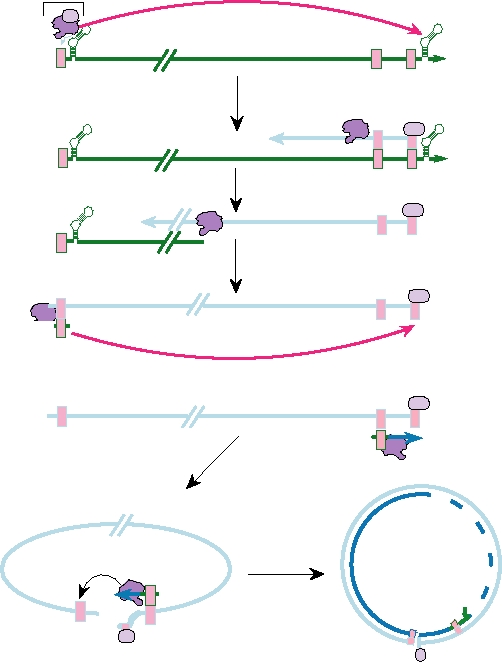
Replication of the Viral Genome
both the 5¢ and 3¢ ends of the RNA, only the 5¢ signal functions
for encapsidation. Once P binds to ε, C is recruited and the cap-
The 3.5-kb pgRNA serves not only as a messenger but also
as an intermediate in viral genome replication. This process
sid assembles. In the absence of P, C assembles into capsids that
is illustrated in Fig. 6.27. Protein P (which has RT activity,
package RNAs randomly. Thus, the specificity in packaging of
including RNase H) uses this RNA as a template to make the
viral RNA lies in the interaction of P with the RNA, unlike most
(partially) dsDNA copy found in the virion. As is the case for
viruses, including the retroviruses, in which it is the capsid pro-
retroviruses, transfer of initiated complexes from one end of
tein that recognizes a packaging signal in the viral genome.
the genome to the other occurs twice during reverse transcrip-
In the nucleocapsid, first-strand synthesis is initiated
tion. Unlike retroviruses, however, the primer for first-strand
by using the -OH group of a specific tyrosine in P as the
synthesis is not a tRNA but protein P itself, which remains
primer. This tyrosine is present in an N-terminal domain
covalently attached to the 5¢ end of the first strand.
(labeled TP in Fig. 6.26) that is distinct from the domain
DNA synthesis takes place in capsids and the first step is there-
that constitutes the RT. Four nucleotides are added, copied
from ε (Fig. 6.27, step 1; see also Fig. 6.28). P with its cov-
fore the encapsidation of the pgRNA. Only pgRNA is packaged.
This RNA is the messenger for both C and P, and both C and
alently attached chain is then transferred to the DR1 accep-
P are required for encapsidation. P binds to a specific sequence
tor site at the 3¢ end of the RNA (which has a sequence
called epsilon present in the 5¢ region of the RNA (Fig. 6.27, step
complementary to the four nucleotides used to start DNA
1). The signal is found within a stem-loop structure present within
synthesis, which are indicated by the red arrow in step (1).
direct repeats of about 200 nt at the two ends of the RNA, which
DNA synthesis of first strand then continues until the 5¢
is illustrated in Fig. 6.28. Interestingly, although ε is present at
end of the RNA is reached (Fig. 6.27, steps 2, 3, and 4).
P
1.
39
59
An pg RNA
CAP 1
2
1
ε
ε
TP
RT
2.
() DNA
2
CAP 1
An pg RNA
2
1
TP
3.
() DNA
RT
2
CAP 1
TP
() DNA
4.
RT 1
2
CAP 1
59
5.
TP
1
() DNA
2
(+) DNA
CAP 1 RT
59
6.
7.
() DNA
(+) DNA
59
RT
CAP
1
59
1
2
() DNA
TP
59
59
FIGURE 6.27 Mechanism of HBV DNA synthesis. Pregenomic RNA (pgRNA) is capped, polyadenylated, and greater
than genome length (green line). It contains two copies each of DR1 and ε, the encapsidation sequence shown in Figure
6.28. Step (1) Priming of reverse transcription occurs when P protein (consisting of TP, RT, and RNaseH domains) makes
a tetranucleotide copy of the bulge in the ε structure. This tetranucleotide is covalently linked to P. The nascent DNA strand
is then translocated to DR1 at the 3′ end of pgRNA. Step (2) TP (terminal protein) is cleaved from P (but see text for an
alternative hypothesis) and remains attached to the 5′ end of the minus strand, while the minus-strand DNA (light blue) is
extended right to left by RT. Step (3) During minus-strand DNA synthesis, RNase H activity degrades pgRNA until P (RT)
reaches the 5′ end of the template RNA. Step (4) A short RNA oligomer is left annealed to a short terminal duplication.
Step (5) The RNA oligomer is translocated to DR2 where it primes plus-strand DNA synthesis (medium blue) left to right.
Step (6) During plus-strand elongation, a second template transfer circularizes the genome and in step (7) the plus strand is
extended a variable length to give mature progeny viral DNA. Modified from Locarnini et al. (1996).
The RNase H activity of P degrades the RNA strand dur-
In the model for DNA synthesis shown in Fig. 6.27, the
ing synthesis of first-strand DNA, but the extreme 5¢ end of
terminal protein (TP) domain is cleaved from the polymerase
the RNA is not degraded. This 5′ piece, which is capped and
domain after initiation of DNA synthesis. However, reports
about 18 nucleotides long, is transferred to the DR2 accep-
conflict as to whether cleavage does or does not occur. If
tor site near the 5′ end of the first-strand cDNA (step 4),
no cleavage occurs, the entire P protein remains covalently
attached to the 5¢ end of the DNA, rather than just TP. If P
where it serves as a primer for second-strand DNA synthesis
(step 5). After reaching the 5′ end of the first-strand cDNA,
remains attached to the 5¢ end of the minus strand, it will
continued synthesis of second-strand cDNA requires trans-
keep the end of the first-strand DNA with it at all times. This
location to the 3′ end of the first strand, as is the case for ret-
could simplify the second jump and the cyclization of the
roviruses (step 6). In the case of hepadnaviruses, cyclization
DNA, since DR2 would be close by.
of the DNA occurs during this translocation, promoted by
Some mature capsids thus produced, which now contain
terminal redundancies. Second-strand synthesis is usually
copies of the viral DNA genome, are transported back into
not complete so that the genomic double-strand DNA has a
the nucleus and release their DNA to amplify the replica-
single-strand gap, of variable length, in it (step 7).
tion cycle. This results in the accumulation in the nucleus
G
CU
U
70
G
C
60
U UC
G
50 U
80
CA
U
G
U
G
C
G
C
G
U
G
C
U
A
U CORE
40 A
C
G
U
G
DR1 20
10
30
U
A
59
39
G
C
CAUGCAACUUUUUCACCUCUGCCUAAUCAUCUCUU
AUUG
PRECORE
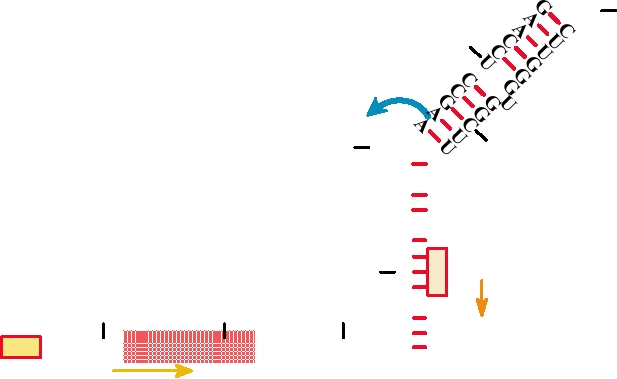
FIGURE 6.28 Two-dimensional structure of the RNA element that forms the packaging signal in hepatitis B RNA.
Numbering is that for the sequence as presented in GenBank. The initiation codons for the precore and core proteins
are marked, the direct repeat sequence (DR1) is shaded, and the blue arrow shows the initiation sequence for DNA
replication (see Figure 6.27). Redrawn from Buckwold and Ou (1999), Figure 2.
of about 20 copies of viral DNA to serve as templates for
of HBsAg contained in these particles can reach more than
100 µg/ml in serum.
mRNA synthesis. This is thought to occur primarily early
in infection, before accumulation of large amounts of S
protein.
HBV and Hepatitis in Humans
HBV has a pronounced tropism for hepatocytes, as do all
Assembly of the Virion
hepadnaviruses, and causes hepatitis in humans. Infection of
Cores, with their partially dsDNA, bud through intra-
neonates or very young children is usually asymptomatic,
cellular membranes to produce mature virions called Dane
but infection nevertheless has serious consequences because
particles. The 42-nm virions contain both L (preS1) and S
chronic infection often results. Infection of adults results in
in their envelopes, with at least one-fourth as much L as
serious disease characterized by liver dysfunction accompa-
S, and both L and S are required for virion assembly. In
nied by jaundice in about one-third of infections, although
the virion, both L and S have both their N and C termini
death from fulminant hepatitis is uncommon (the fatality
outside. Thus, they must span the membrane at least twice,
rate is around 1% of acute cases). The incubation period is
and some models propose that they span the membrane four
long, 45120 days, and convalesence is usually extended
times. L is myristoylated at the N terminus. Myristoylation
(more than 2 months), but more than 90% of adults infected
is required for the infectivity of the virus but not for assem-
by HBV recover completely. In the United States, almost
bly, suggesting that myristoylation serves a function in
500,000 cases of hepatitis are reported annually, of which
entry. Virions also contain M in amounts equivalent to L,
about 40% are due to HBV (Table 3.6). It is reasonable to
but M does not appear to be required for assembly or for
assume that the total number of new infections with HBV
infectivity of virions.
is perhaps 10-fold the number of reported cases of HBV-
In addition to the 42-nm Dane particles, 20-nm particles
induced hepatitis.
are also produced in abundance (104- to 106-fold excess over
In a small number of adult cases, less than 10%, the infec-
Dane particles). These particles contain S and M but little or
tion is not cleared and becomes chronic. Infection of neonates
no L, and lack the core. These particles form when S alone
or young children results in high levels of chronicity, how-
is expressed in cells, and thus S has the ability to produce a
ever. Up to 90% of neonates infected with HBV become
bud in the absence of other viral components. The virus may
chronically infected, and infection of 3-year-old children
produce such vast quantities of the 20-nm particles, which
may result in 30% chronicity. It is thought that the immature
can result in concentrations as high as 1013/ml in serum, in
state of the immune system in the very young is important
order to tolerize the immune system, since the concentration
in the development of a chronic infection. Chronic infection
to 90% of babies born to mothers who are acutely or chroni-
may remain asymptomatic and may even eventually clear
cally infected with HBV and positive for HBeAg will be
in a small fraction of cases, especially if infection occurred
infected by HBV, and most of these will become chronically
as an adult. However, other patients develop chronic active
infected. In the United States, there are an estimated 1.2 mil-
hepatitis that may progress to cirrhosis and death.
lion carriers of HBV, and worldwide there are an estimated
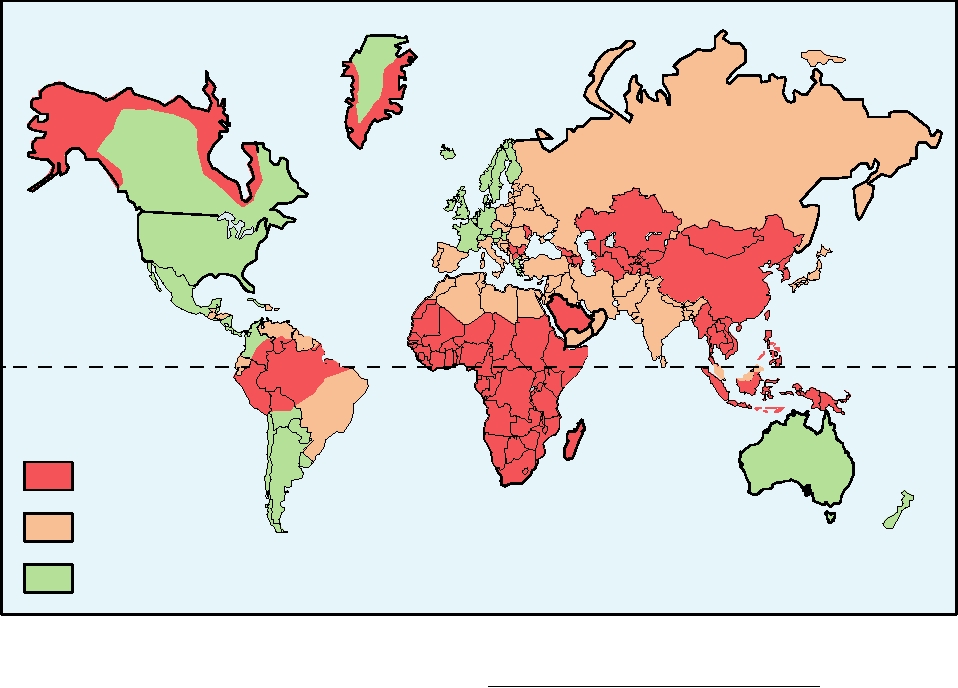
HBV is spread primarily by contact with contaminated
350 million. The fraction of the population chronically
blood, by sexual intercourse, and from mother to child dur-
infected with HBV varies from 0.1 to 0.5% in developed
ing delivery or breast-feeding. Persistently infected indi-
countries to 5 to 15% in Southeast Asia and sub-Saharan
viduals can have very high titers of virus in the blood, up to
1010/ml, and the virus resists drying for up to 1 week. Thus,
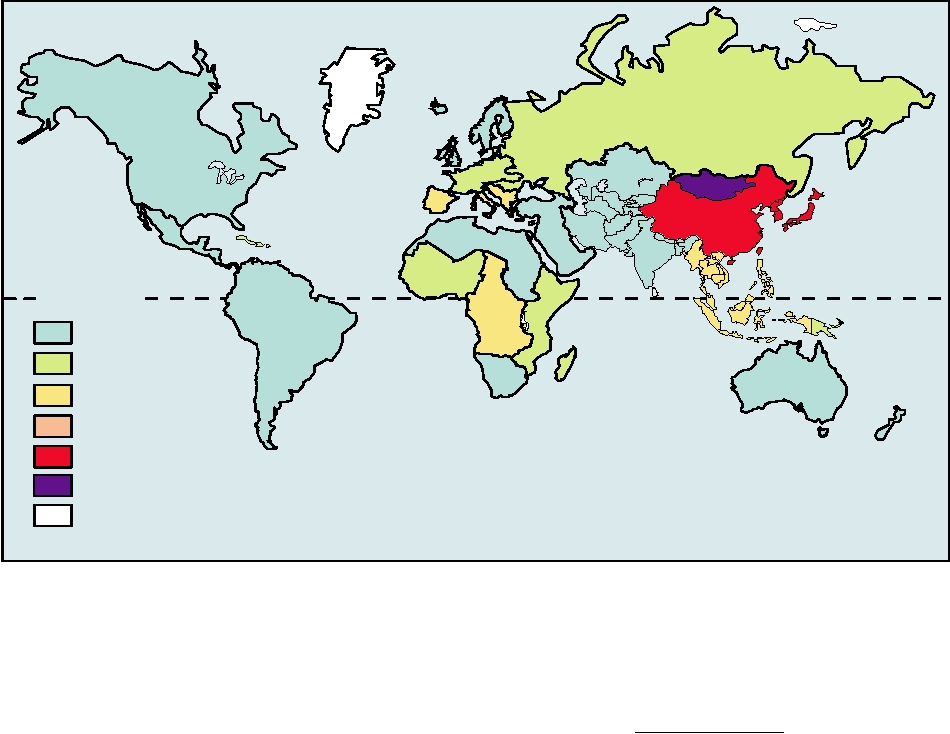
Africa. A map that illustrates the prevalence of HBV in
contact with infected blood need not be extensive to transmit
different regions of the world is shown in Fig. 6.29.
the virus. It has been suggested that household contact leads
to spread via sharing of razors, for example. Medical person-
HBV and Hepatocellular Carcinoma
nel are at risk of contracting the virus from their patients, not
only by needle stick, which is responsible for many cases
Liver cancer causes more than 500,000 deaths a year
in unvaccinated individuals, but through other contact with
worldwide, and about 90% of primary malignant tumors of
contaminated blood. The virus also spreads readily among
the liver are hepatocellular carcinoma (HCC). HCC is more
institutionalized individuals. At one time, blood transfusion
common in men than women, by 4 to 1, and is in the top 10
was a source of spread of virus, but with the development
in frequency of cancers in humans. HCC is more common
of sensitive assays for the presence of the virus, the risk of
in regions that exhibit high chronicity for HBV (compare
infection following blood transfusion in developed countries
Fig. 6.30 with Fig. 6.29). The association of HBV with HCC
is now 1/200,000 per unit of blood.
is also shown by data such as the finding that in areas in
Chronic infection acquired at birth is thought to be the
which chronic infection occurs in 510% of the population,
major mechanism by which the virus persists in nature. Up
5080% of HCC patients are chronically infected with HBV.
Equator
Endemicity
High (>8% carriers,
50-95% seropositive)
Medium (2-7% carriers,
30-50% seropositive)
Low (<2% carriers,
few seropositives)
FIGURE 6.29 Worldwide hepatitis B prevalence as of 2005. Each country is designated as having high, medium, or
low levels of endemicity, based on estimates of carrier frequency. Correlated with this is a range of persons who show
serological evidence of past infection. From CDC Web page: http://www.cdc.gov/ncidod/diseases/hepatitis/slides.
17
18
9
20
15
13
19
11
12
10
7
6
16
3
5
14
Equator
Incidence
2
Cases/100,000
22
1
8
0- 5.0
5.1-10.0
4
21
10.1-15.0
15.1-20.0
20.1-40.0
> 40.1
No Data
FIGURE 6.30 Average annual incidence (cases per 100,000) of liver cancer in various regions of the world in 2002. Of
these cases, 67% are due to chronic hepatitis B and 26% are due to chronic hepatitis C infection. The names of the regions
and the incidence for men/women are (1) Eastern Africa 10.7/4.8; (2) Middle Africa 15.4/9.0; (3) Northern Africa 2.6/1.6; (4)
Southern Africa 4.3/1.8; (5) Western Africa 9.0/3.5; (6) Caribbean 7.5/4.7; (7) Central America 3.2/3.6; (8) South America
2.9/2.6; (9) North America 7.0/3.2; (10) China 37.9/15.1; (11) Japan 43.8/19.2; (12) North and South Korea 45.7/13.0;
(13) Mongolia 53.1/34.2; (14) Southeast Asia 13.2/4.6; (15) South Central Asia 1.8/1.1; (16) Western Asia 3.0/1.5; (17)
Eastern Europe 6.8/4.3; (18) Northern Europe 5.41/3.4; (19) Southern Europe 19.8/8.8; (20) Western Europe 10.1/3.6; (21)
Australia/New Zealand 5.4/2.0; (22) Melanesia 10.2/5.5. Data from Globocan 2002 at: http: //www-dep.iarc.fr/.
The lifetime risk of HCC in chronically infected persons is
evidence that the X protein might be responsible, at least in
estimated to be 1025%. It is clear that chronic HBV infec-
part, for HCC caused by HBV. The HBV X gene can induce
tion contributes to a large fraction of HCC cases.
HCC in transgenic mice. This protein binds to p53, a known
The mechanism by which chronic infection by HBV leads
anti-oncogene (Chapter 7) that regulates signaling pathways
to HCC is not altogether clear, and may not be the same in
and modifies the activities of transcription factors. Although
all cases. One possibility is that long-term infection, charac-
it is probable that the X gene product is responsible for
terized by continuing destruction of liver cells followed by
induction of HCC in some fraction of cases, it cannot be the
regrowth, results eventually in the appearance and selection of
whole story because chronic infection by hepatitis C virus
tumor cells. Up to 90% of patients with HCC associated with
(Chapter 3) also leads to HCC, and HCV lacks the X gene.
HBV infection have cirrhosis, implying extensive liver dam-
age. It also appears that HCC may result from other causes
The Immune System and HBV
of liver disease such as alcohol-induced cirrhosis or chronic
infection by HCV (see Chapter 3). Furthermore, HCC often
HBV infection of itself does not lead to the death of
appears only after 3040 years of chronic infection by HBV.
infected hepatocytes. Whether in vivo or in cell culture, a
Thus, there is an association between HCC and continuing
persistent, noncytolytic infection is established by the virus.
liver damage and regeneration over very long periods.
Liver damage during HBV infection results instead from the
Chronic infection of woodchucks by WHV results in
activities of cytotoxic T lymphocytes (CTLs) (see Chapter
HCC, and in 40% of HCC cases in this system there is inte-
10), which attempt to clear the infection by killing infected
gration of the WHV DNA genome near N-myc2. There is
cells. It appears that the strength of the CTL response deter-
no evidence in humans that insertional mutagenesis of HBV
mines the course of infection. A vigorous response results in
DNA is responsible for HCC in humans, but there is some
clearance and recovery, although often after frank hepatitis
with jaundice. A weak response results in chronic infection
of continuing treatment. Adefovir dipivoxil, an analogue of
with little symptomology. An intermediate response results
adenosine, is also effective in controlling virus replication
in chronic infection characterized by chronic hepatitis.
in about half of patients tested but most patients relapse
Because of the potential seriousness of chronic infection
when therapy is terminated. Thus, there is as yet no therapy
by HBV, including the potential to infect others, continuing
that is effective in clearing virus infection in even half of
efforts are being made to develop methods of controlling or
patients.
clearing the infection in chronically infected people. HBeAg
Liver transplantation is offered to some patients with
was used as a marker of severity of infection until recently.
HBV infection. However, circulating virus invariably rein-
This has now been replaced clinically by direct measure-
fects the graft.
ment of viral titers, which is used to assess the response to
therapy.
Vaccination against HBV
The first treatment that showed at least partial success
Several vaccines have been developed to prevent infec-
was use of high doses of interferon for extended periods of
tion by HBV. The first vaccine, which was licensed in 1981,
time. This succeeds in clearing the viral infection in a small
was prepared from blood plasma from chronically infected
minority of patients. Interferon is a cytokine that boosts the
individuals. It consisted of highly purified preparations of
immune response (see Chapter 10), and these doses of inter-
20-nm particles that were treated to inactivate any residual
feron appear to enable the immune system to eradicate the
virus infectivity (whether HBV or any other virus). This vac-
virus in those patients that respond. However, the drug is
cine was effective and safe, but obvious difficulties accom-
poorly tolerated, with significant side effects. Nucleoside
pany the production of large amounts of such a vaccine.
analogues have also been tested. Most are not effective but
Recombinant vaccines in which the S gene is expressed in
lamivudine treatment resulted in improvement in more than
yeast or in Chinese hamster ovary cells have now replaced
50% of patients in a large trial, and the apparent clearing of
this early vaccine. These vaccines are cheaper and can be
infection in 16% of patients. Resistant viruses appeared in
produced in large quantities.
more than 10% of cases, however, limiting the effectiveness
Equator
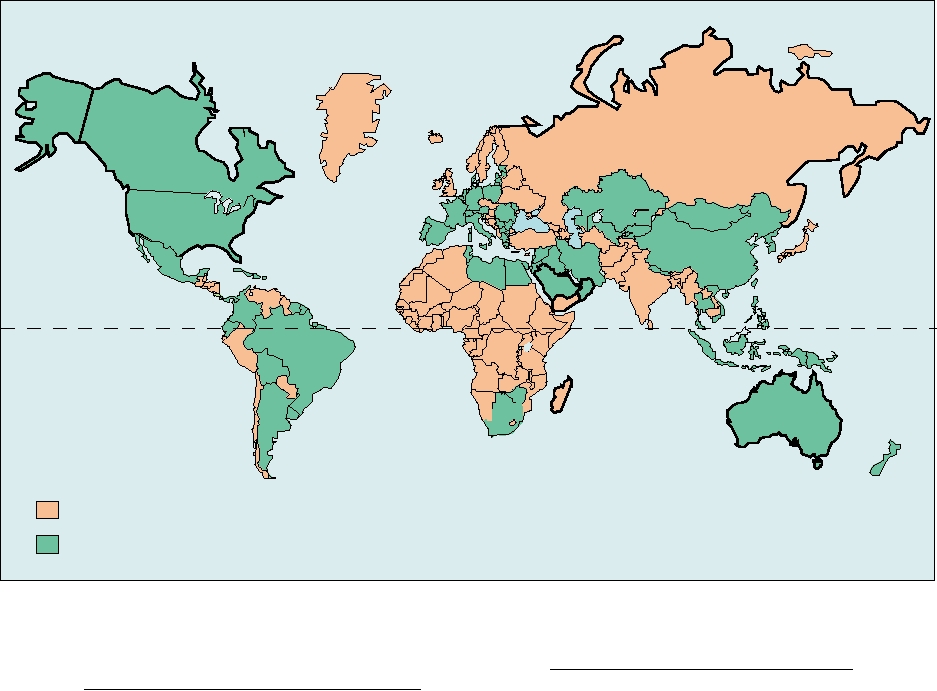
HepB3 vaccine coverage zero
HepB3 vaccine coverage >50%
FIGURE 6.31 Global status of hepatitis B immunization policy, as of October 2005. Countries that have policies in
place for routine immunization of infants (as measured by estimates of coverage for the third dose of hepatitis B vaccine:
HepB3). Countries whose reported HepB3 coverage is zero are presumed not to have such programs in place. From WHO
Program on Immunization, Surveillance, Assessment, and Monitoring at: http://www.who.int/immunization_monitoring/
en/globalsummary/timeseries/tscoveragehepb3.htm.
Search WWH :